Green algae. Order Green algae: brief description What pigment is contained in red algae
The division of algae into systematic groups - divisions - basically coincides with the nature of their color, associated, of course, with structural features. The most widespread division of algae into 10 divisions: 1) blue-green, 2) pyrophytic, 3) golden, 4) diatoms, 5) yellow-green, 6) brown, 7) red, 8) euglenophytes, 9) green, 10) characeae.
Blue-green algae. They are colored bluish-green, sometimes almost black-green or olive green. Pigments: chlorophyll A, carotenoids, blue phycocyan, and a small amount of red phycoerythrin. The forms are predominantly multicellular, colonial or filamentous, some are unicellular. The shells consist of murein, pectin substances, and sometimes cellulose, and mucus. Characteristic structure of cells: there is no differentiated nucleus, chloroplasts, vacuoles; photosynthetic membranes, pigments and nucleoproteins are located in the cytoplasm - the main component of the nuclei of other plants. Many blue-green algae have a network of gas vacuoles in the cytoplasm; a significant number of filamentous algae form so-called heterocysts - cells with a specific structure.
Reproduction in unicellular organisms occurs by cell division, in colonial and filamentous organisms - by the disintegration of colonies and filaments, and there is a sexual process. Many blue-greens produce spores to survive unfavorable conditions and for reproduction.
Pyrophytic algae are unicellular; an essential feature is the dorsoventral (dorsoventral) structure of their cells (the dorsal, ventral and lateral sides, the anterior and posterior ends are clearly expressed). The presence of grooves is characteristic; there can be two of them (longitudinal and transverse) or one (longitudinal). There are two flagella of different lengths, a pharynx (which looks like a bag, a tube, an internal pocket or a triangular reservoir), and strongly refracting light bodies - trichodists (located in the peripheral layer of the cytoplasm, on the inner surface of the pharynx or inside the protoplast). They are usually colored olive, brown or brown, often yellow, golden, red, less often blue, indigo. Pigments: chlorophylls a and c, xanthophylls, peridinins. There are colorless forms. Nutrition is autotrophic, less often heterotrophic. Reproduction is mainly vegetative, less often asexual (zoo- and autospores). The reproductive process is unknown. Widely distributed in water bodies of our planet (fresh, brackish waters, seas).
Golden algae. Mostly microscopic, unicellular, colonial and multicellular forms. Painted golden yellow. Pigments: chlorophylls A and c, carotenoids, especially a lot of fucoxanthin. They live mainly in clean fresh waters, characteristic of the acidic waters of sphagnum bogs. Some species live in the seas. They usually develop in early spring, late autumn and winter. The structure of the cell is the same: the protoplast contains one or two wall trough-shaped chloroplasts with a pyrenoid, the nucleus is small, in some species there are one or two pulsating vacuoles at the front of the cell. Shell; in the simplest representatives - a delicate periplast; in most golden ones, the peri-layer is dense, the cell has a constant shape; highly organized representatives have a real cellulose, usually double-circuit shell. In many species, the cells are covered with a shell with spines and spines. They reproduce by simple cell division or disintegration of the thallus into parts. Asexual reproduction is observed with the help of zoospores, less often autospores. The sexual process is known in the form of typical isogamy, autogamy.
Diatoms. Microscopic unicellular, colonial or filamentous algae of brownish-yellow color. Pigments: chlorophylls A and with carotene, fucoxanthin and other xanthophylls. The shape of the cells is varied. The shell is represented by a shell of silica, lined from the inside with a pectin layer. The flint shell contains pores - areoles. Depending on the configuration of the cells and the structure of the valves, diatoms with radial and bilateral symmetry are distinguished. Many diatoms have a longitudinal slit in their valves (the so-called suture). At its ends and in the middle there are thickenings of the shell, which are called nodules. Thanks to the seam and knots, the cell moves. Algae that do not have a seam do not move.
Protoplasm is located in cells in a thin layer. The cell contains a nucleus, a vacuole with cell sap. Chloroplasts have different shapes; there are one or several of them.
Diatoms reproduce by division. They also have a sexual process, usually associated with the formation of auxospores, i.e. “growing coop", which grow strongly and then grow into cells that differ significantly in size from the original ones. Auxospores are characteristic only of diatoms. Diatoms can form resting spores.
They live everywhere: in bodies of water (fresh and salty), in swamps, on stones and rocks, in soils and on their surface, on snow and ice.
Yellow-green algae. Unicellular, colonial, multicellular and noncellular forms. Most are immobile, but there are mobile forms.
This division has recently been isolated from green algae. Yellow-green algae are distinguished by the fact that the two flagella of their zoospores are unequal in location and structure: one is longer, directed forward, has processes on its axis, the second is smooth, short, directed backward. The cell membrane contains a lot of pectin substances and does not react to cellulose.
The cell structure of yellow-green algae is the same. The protoplast contains several chloroplasts that have a disc-shaped, trough-shaped, lamellar, less often ribbon-shaped, stellate shape. Main pigments: chlorophyll a, e, carotenes and xanthophylls. In motile forms, a red eye is located at the anterior end of the chloroplast. There is one nucleus. In a few species there is a pyrenoid and in the anterior part of the clerk there are one or two pulsating vacuoles. Vegetative reproduction occurs by cell division, disintegration of colonies or filaments, asexual reproduction occurs by zoo- and autospores. The sexual process is known for a small number of genera: iso-, oogamy. Distributed in plankton, benthos of fresh water bodies, in the seas, soil, and in places with high humidity.
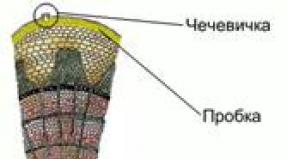
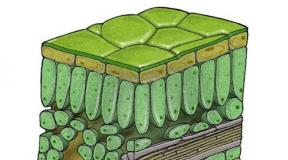
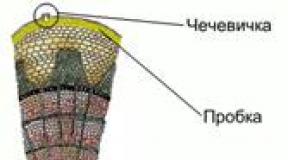
Brown algae. In most cases these are marine forms. They are multicellular, attached to the substrate. Their sizes vary from a few millimeters to several meters, there are species reaching 60 m. In appearance, they are branched bushes, plates, cords, ribbons, some seem to have a stem and leaves. The cell has one nucleus, chloroplasts are brown, granular, and often there are many of them. Pigments: chlorophylls A And With, carotenes, a lot of fucoxanthins. In multirow thalli, specialization of cells with the formation of tissues is observed. In the simplest case, a distinction is made between the cortex (intensely colored cells containing chloroplasts) and the pith (colorless large cells of the same shape). In more complexly organized ones (kelp and fucus) there is also a superficial layer of dividing cells capable of producing reproductive organs and called meristoderm, and an intermediate layer between the core and the cortex. The core serves for the transport of photosynthetic products and performs a mechanical function.
Red algae (purple algae). An extensive group among bottom seaweeds. Very rarely found in fresh water bodies (batrachospermum species) and in terrestrial fouling (porphyridium). Colored in various shades of red, some are yellowish, olive or bluish-green in color. Pigments: chlorophylls aAnd d (the latter is found only in red algae), carotenes, xanthophylls, R-phycoerythrin, R-phycocyanin. Almost all red algae are multicellular, in the form of filaments, branched filaments, bushes, and some have stem-like and leaf-like organs. They don't come in sizes as big as brown ones. All are attached to the substrate. The cells are covered with a shell consisting of two layers: the inner one is cellulose and the outer one is pectin. The most simply organized cells are mononuclear, while the highly organized ones are multinucleated. There are one or more chloroplasts. A feature of red algae is the presence of special glandular cells in some representatives. The cells that make up the thallus are connected by pores.
Vegetative propagation is rare. In an asexual process, zoospores are completely absent. The sexual process is oogamy.
Purpleworts have a unique structure of the oogonium (carpogon) and complex processes of zygote development. There are no moving stages in the development cycle. The zygote, before giving rise to the sporophyte, undergoes a complex development, as a result of which spores (carpospores) are formed, giving rise to the sporophyte,
Euglena algae. Microscopic organisms. The shape of the cells is mainly elliptical and spindle-shaped. Chloroplast is stellate, ribbon-shaped, large-lamellar in shape. Pigments: chlorophylls a, b , carotene, xanthophylls. Some euglenoids have a red pigment, astaxanthin, which regulates the amount of light reaching the chloroplasts. Under intense lighting conditions, the pigment accumulates in the peripheral part of the cell and obscures the chloroplasts. The cell turns red. There is no cellulose membrane; its role is played by a compacted layer of cytoplasm; some have a shell that is not tightly connected to the protoplast. At the anterior end of the body there is a depression (pharynx), from the bottom of which one or two flagella extend. Euglenaceae move by changing the shape of the body and with the help of a flagellum, simultaneously turning around the longitudinal axis. In living euglenoids, in the front part of the cell there is a red spot - the stigma, which acts as a light-sensitive organ. They reproduce by longitudinal division. The presence of sexual process has not been established. They live mainly in small fresh water bodies, some in brackish ones.
Charovaya algae. A peculiar group of algae, in appearance reminiscent of higher plants. Widely distributed in freshwater ponds and lakes, especially in hard, calcareous water; found in sea bays and brackish continental waters. They form thickets. The height of their thalli is usually 20 - 30 cm, but can reach 1 and even 2 m. They have the appearance of bushy thread-like or stem-like green shoots of an articular-whorled structure: on the main shoots, conventionally called stems, whorls of lateral shoots are located at some distance from each other - conditionally leaves are also segmented in structure. The locations of the whorls are called nodes, and the sections of the stem between them are called internodes. The cells of nodes and internodes are different: an internode is a giant elongated cell that is not capable of division; the node consists of several small mononuclear cells collected in a disk, differentiating during the process of division and forming lateral branches and a whorl. Chloroplasts are numerous and have the shape of small disc-shaped bodies (resembling chlorophyll grains). Pigments: chlorophylls A and b , carotenes, xanthophylls (similar to green algae). The structure of the sexual reproductive organs formed at the top of most segments - leaves - is peculiar. Female organs - oogonia - and male organs - antheridia - are multicellular, usually developing on the same plant (rarely dioecious).
Source---
Bogdanova, T.L. Handbook of biology / T.L. Bogdanov [and others]. – K.: Naukova Dumka, 1985.- 585 p.
Far Eastern State Technical
fisheries university
Institute of Marine Biology named after. A.V. Zhirmunsky Far Eastern Branch of the Russian Academy of Sciences
L.L. Arbuzova
I.R. Levenets
Seaweed

Reviewers:
– V.G. Chavtur, Doctor of Biological Sciences, Professor of the Department of Marine Biology and Aquaculture of the Far Eastern State University
– S.V. Nesterova, Ph.D., senior researcher at the Laboratory of Flora of the Far East, Botanical Garden-Institute, Far Eastern Branch of the Russian Academy of Sciences
Arbuzova L.L., Levenets I.R. Algae: Study. village Vladivostok: Dalrybvtuz, IBM FEB RAS, 2010. 177 p.
The manual provides modern information about the anatomy, morphology, taxonomy, lifestyle and practical significance of algae.
The textbook is intended for bachelor students in the areas of “Aquatic Bioresources and Aquaculture” and “Ecology and Environmental Management” full-time and part-time, masters of ecology, biology, ichthyology and fish farming.
©Far Eastern State
technical fishery
university, 2010
©Institute of Marine Biology named after. A.V. Zhirmunsky Far Eastern Branch of the Russian Academy of Sciences, 2010
ISBN ……………………..
Introduction…………………………………………………………………………………
1. Structure of algae cells……………………………………
2. General characteristics of algae……………………………
2.1. Types of food………………………………………………………
2.2. Types of thalli…………………………………………………..
2.3. Reproduction of algae……………………………………..
2.4. Life cycles of algae……………………………….
3. Ecological groups of algae…………………………….
3.1. Algae of aquatic habitats…………………………..
3.1.1. Phytoplankton………………………………………………….
3.1.2. Phytobenthos……………………………………………………..
3.1.3. Algae of extreme aquatic ecosystems……………
3.2. Algae of non-aquatic habitats………………………
3.2.1. Aerophilic algae…………………………………….
3.2.2. Edaphilic algae…………………………………….
3.2.3. Lithophilic algae……………………………………..
4. The role of algae in nature and practical significance………
5. Modern taxonomy of algae………………………..
5.1. Prokaryotic algae………………………………..
5.1.1. Department Blue-green algae………………………
5.2. Eukaryotic algae…………………………………….
5.2.1. Department Red algae…………………………….
5.2.2. Section Diatoms………………………..
5.2.3. Department Heterokont algae…………………….
Class Brown algae …………………………………
Class Golden algae …………………………
Class Sinura algae …………………………..
Class Pheotamnia algae………………………
Class Raphid algae ………………………….
Class Eustigma algae …………………………
Class Yellow-green algae………………………
5.2.4. Department Prymnesiophyte algae……………….
5.2.5. Department Cryptophyte algae……………………
5.2.6. Department Green algae…………………….………
5.2.7. Division of Characeae…………………………….
5.2.8. Department Dinophyte algae………………………
5.2.9. Section Euglena algae …………………………
Literature………………………………………………………………………………
Glossary of terms……………………………………………………….
Application …………………………………………………………….
INTRODUCTION
Algae traditionally comprise a diverse group of thallus, photosynthetic, spore-bearing, avascular organisms. Like all lower plants, the reproductive organs of algae are devoid of integument, the body is not divided into organs, and there are no tissues. Among algae there are both eukaryotic and prokaryotic forms. The latter, unlike chlorobacteria, release free oxygen into the environment during photosynthesis.
Algae occupy a dominant position in both fresh and marine water bodies. Being the main producers, they largely determine the fish productivity of aquatic ecosystems. Thanks to photosynthetic activity, algae enrich the water with oxygen and reduce the amount of carbon dioxide. They have a unique ability to accumulate various harmful substances from the surrounding aquatic environment, as well as release metabolites into the environment that suppress the growth of pathogenic microorganisms. Algae, by changing the chemical composition of water, often contribute to its purification. The qualitative and quantitative composition of algae groups is an important indicator of the ecological state of water bodies. A number of species are used as indicators of aquatic pollution.
The study of algae is an important stage in the training of specialists in the field of mariculture, fish farming and marine ecology. Knowledge of the structure, ecology and systematics of algae is basic for the study of hygrobiology, ichthyology, ecology, ichthyotoxicology; they are also necessary for assessing the raw material base of reservoirs and drawing up fishing forecasts.
Recently, thanks to modern methodological techniques, new information has been obtained about the fine structure, physiology and biochemistry of algae, which has caused a revision of traditional ideas. The taxonomy of lower plants, which include algae, has undergone the greatest changes. At the same time, modern information about the systematics and structure of algae is not reflected in educational literature on botany, and special literature on phycology is not available to a wide student audience.
This textbook provides the latest information about the structure, morphology, taxonomy, ecology and practical significance of algae. A description of the most significant algae taxa is given.
The textbook is intended for bachelor students in the areas of “Aquatic Bioresources and Aquaculture” and “Ecology and Environmental Management” of full-time and part-time forms of study, masters in the field of ecology, ichthyology, fish farming and aquaculture.
Teachers from Dalrybvtuz and specialists in the field of hydrobiology and phycology of the Far Eastern Branch of the Russian Academy of Sciences took part in the preparation of materials for this manual.
1. STRUCTURE OF ALGAE CELLS
Prokaryotic algae are similar in cell structure to bacteria: they lack membrane organelles, such as a nucleus, chloroplasts, mitochondria, endoplasmic reticulum, and Golgi apparatus.
Eukaryotic algae contain structural elements characteristic of higher plant cells (Fig. 1).

Rice. 1. An adult plant cell without secondary wall thickening (schematized) at maximum magnification of a light microscope (by: 1 – cell wall, 2 – median plate, 3 – intercellular space, 4 – plasmodesmata, 5 – plasmalemma, 6 – tonoplast, 7 – central vacuole, 9 - nucleus, 10 - nuclear envelope, 11 - pore in the nuclear envelope, 12 - nucleolus, 13 - chromatin, 14 - chloroplast, 15 - grana in the chloroplast, 16 - starch grain in the chloroplast, 17 - mitochondrion, 18 - dictyosome, 19 – granular endoplasmic reticulum, 20 – droplet of reserve fat (lipid) in the cytoplasm, 21 – microbody, 22 – cytoplasm (hyaloplasm)
The above diagram of a plant cell generally reflects the structure of algae cells, however, many algae, along with typical plant organelles (plastids, vacuole with cell sap), contain structures characteristic of animal cells (flagella, stigma, membranes atypical for plant cells).
Cell covers
Cell covers ensure the resistance of the internal contents of cells to external influences and give the cells a certain shape. The covers are permeable to water and low molecular weight substances dissolved in it and easily transmit sunlight. The cell covers of algae are distinguished by great morphological and chemical diversity. They include polysaccharides, proteins, glycoproteins, mineral salts, pigments, lipids, and water. Unlike higher plants, there is no lignin in the shells of algae.
The structure of cell covers is based on the plasmalemma, or cytoplasmic membrane. In many flagellar and amoeboid representatives, the cells on the outside are covered only with plasmalemma, which is not capable of providing a constant body shape. Such cells can form pseudopodia. Based on morphology, several types of pseudopodia are distinguished. Most often found in algae rhizopodia, which are thread-like long, thin, branched, sometimes anastomosing cytoplasmic projections. There are microfilaments inside the rhizopodia. Lobopodia– wide rounded protrusions of the cytoplasm. They are found in algae with amoeboid and monadic types of thallus differentiation. Less common in algae filopodia- thin mobile formations resembling tentacles that can be retracted into the cell.
Many dinoflagellates have bodies covered with scales located on the surface of the cell. The scales can be single or close into a continuous cover - flowing. They may be organic or inorganic. Organic scales are found on the surface of green, golden, cryptophyte algae. The composition of inorganic flakes may include either calcium carbonate or silica. Calcium carbonate flakes – coccoliths– found predominantly in marine primnesiophyte algae.
Often, the cells of flagellated and amoeboid algae are located in houses that are mainly of organic origin. Their walls can be thin and transparent (genus Dinobryon) or more durable and colored due to the deposition of iron and manganese salts in them (genus Trachelomonas). The houses usually have one hole for the flagellum to exit, sometimes there may be several holes. When algae multiply, the house is not destroyed; most often, one of the resulting cells leaves it and builds a new house.
The cell covering of euglenoid algae is called a pellicle. The pellicle is a collection of the cytoplasmic membrane and the protein bands, microtubules and cisterns of the endoplasmic reticulum located under it.
In dinophyte algae, the cell covers are represented by amphiesma. Amphiesma consists of a plasmalemma and a set of flattened vesicles located under it, under which lies a layer of microtubules. The vesicles of a number of dinophytes may contain cellulose plates; this is called amphiesma current, or shell(birth Ceratium, Peridinium).
In diatoms, a special cell cover is formed on top of the plasmalemma - shell, consisting mainly of amorphous silica. In addition to silica, the shell contains an admixture of organic compounds and some metals (iron, aluminum, magnesium).
In the cell walls of green, yellow-green, red and brown algae, the main structural component is cellulose, which forms a framework (structural basis) immersed in a matrix (semi-liquid medium) consisting of pectin, hemicellulose, alginic acid and other organic substances.
Flagella
Monadic vegetative cells and monadic stages in the life cycle (zoospores and gametes) of algae are equipped with flagella - long and rather thick cell outgrowths, externally covered with plasmalemma. Their number, length, morphology, place of attachment, and pattern of movement are quite diverse in algae, but are constant within related groups.
The flagella may be attached at the anterior end of the cell (apical) or may be slightly moved to the side (subapical); they can be attached to the side of the cell (laterally) and on the ventral side of the cell (ventrally). Flagella identical in morphology are called isomorphic, if they differ - heteromorphic. Isocont- these are flagella of the same length, heterokontnye– different lengths.
Flagella have a single structure plan. One can distinguish the free part (undulipodium), transition zone, and basal body (kinetosome). Different parts of the flagellum differ in the number and arrangement of microtubules, which form the skeleton (Fig. 2).

Rice. 2. Scheme of the structure of algae flagella (according to: L.L. Velikanov et al., 1981): 1 – longitudinal section of the flagella; 2, 3 – transverse section through the tip of the flagellum; 4 – transverse section through the undulipodium; 5 – transition zone; 6 – cross section through the base of the flagellum – kinetosome
Undulipodium(translated from Latin as “wavepod”) is capable of making rhythmic wave-like movements. An undulipodium is a membrane-clad axoneme. Axoneme consists of nine pairs of microtubules arranged in a circle and a pair of microtubules in the center (Fig. 2). Flagella can be smooth or covered with scales or mastigonemes (hairs), and in dinophytes and cryptophytes they are covered with both scales and hairs. The flagella of prymnesiophyte, cryptophyte and green algae can be covered with scales of various shapes and sizes.
Transition zone. Functionally, it plays a role in strengthening the flagellum at the site of its exit from the cell. In algae, there are several types of transition zone structures: transverse plate (dinophytes), star-shaped structure (green), transition spiral (heterocontine), transition cylinder (primnesiophytes and dinophytes).
Basal body or kinetosome. This part of the flagellum has a structure in the form of a hollow cylinder, the wall of which is formed by nine triplets of microtubules. The function of the kinetosome is the connection of the flagellum with the plasmalemma of the cell. The basal bodies of a number of algae can take part in nuclear division and become centers of microtubule organization.
Mitochondria
Mitochondria are found in eukaryotic algae cells. The shape and structure of mitochondria in algae cells are more diverse compared to mitochondria of higher plants. They can be round, thread-like, network-shaped or irregular in shape. Their shape can vary in the same cell at different stages of the life cycle. Mitochondria are covered with a shell of two membranes. The mitochondrial matrix contains ribosomes and mitochondrial DNA. The inner membrane forms folds - cristas(Fig. 3).

Rice. 3. Structure of plant mitochondria (according to:): A – volumetric image; B—longitudinal section; B – part of the crista with mushroom-shaped protrusions: 1 – outer membrane, 2 – inner membrane, 3 – crista, 4 – matrix, 5 – intermembrane space, 6 – mitochondrial ribosomes, 7 – granule, 8 – mitochondrial DNA, 9 – ATP-somes
Algae cristae come in various shapes: disc-shaped (Euglenoid algae), tubular (Dinophyte algae), lamellar (green, red, cryptomonad algae) (Fig. 4).

Rice. 4. Different types of mitochondrial cristae (according to:): A – lamellar; B – tubular; B – disc-shaped; k - cristae
Disc-shaped cristae are considered the most primitive.
Pigments
All algae differ well in their set of photosynthetic pigments. Such groups in plant taxonomy have the status of divisions.
The main pigment of all algae is the green pigment chlorophyll. There are four known types of chlorophyll, which differ in their structure: chlorophylla– present in all algae and higher plants; chlorophyll b– found in green, charophyte, euglenoid algae and higher plants: plants containing this chlorophyll always have a bright green color; chlorophyll c– found in heterokont algae; chlorophyll d– a rare form, found in red and blue-green algae. Most photosynthetic plants contain two different chlorophylls, one of which is always chlorophyll a. In some cases, instead of the second chlorophyll, there are biliproteins. There are two types of biliproteins found in blue-green and red algae: phycocyanin– blue pigment, phycoerythrin- red pigment.
The obligatory pigments included in photosynthetic membranes are yellow pigments - carotenoids. They differ from chlorophylls in the spectrum of absorbed light and are believed to perform a protective function, protecting chlorophyll molecules from the destructive effects of molecular oxygen.
In addition to the listed pigments, algae also contain: fucoxanthin– golden pigment; xanthophyll- brown pigment.
Plastids
Pigments in the cells of eukaryotic algae are located in plastids, as in all plants. There are two types of plastids in algae: colored chloroplasts (chromatophores) and colorless leucoplasts (amyloplasts). The chloroplasts of algae, in contrast to those of higher plants, are much more diverse in shape and structure (Fig. 5).

Rice. 5. Scheme of the structure of chloroplasts in eukaryotic algae (by:): 1 – ribosomes; 2 – chloroplast shell; 3 – girdle thylakoid; 4 – DNA; 5 – phycobilisomes; 6 – starch; 7 – two membranes of chloroplast EPS; 8 – two membranes of the chloroplast shell; 9 – lamella; 10 – spare product; 11 – core; 12 – one membrane of chloroplast EPS; 13 – lipid; 14 – grain; 15 – pyrenoid. A – thylakoids are located one at a time, there is no CES - chloroplast endoplasmic reticulum (Rhodophyta); B – two-thylakoid lamellae, two CES membranes (Cryptophyta); B – trithylakoid lamellae, one CES membrane (Dinophyta. Euglenophyta); D – trithylakoid lamellae, two CES membranes (Heterokontophyta, Prymnesiophyta); D – two-, six-thylakoid lamellae, no CES (Chlorophyta)
The structural photosynthetic unit of eukaryotes and prokaryotes is thylakoid- flat membrane sac. Thylakoid membranes contain pigment systems and electron carriers. The light phase of photosynthesis is associated with thylakoids. The dark phase of photosynthesis takes place in the stroma of the chloroplast. The shell of green and red algae consists of two membranes. In other algae, the chloroplast is surrounded by an additional one or two membranes of the chloroplast endoplasmic reticulum(HES). In euglenaceae and most dinophytes, the chloroplast is surrounded by three membranes, and in heterokontaceae and cryptophytes - by four (Fig. 5).
Nucleus and mitotic apparatus
The algae nucleus has a structure typical of eukaryotes. The number of nuclei in a cell can vary from one to several. On the outside, the nucleus is covered with a shell consisting of two membranes; the outer membrane is covered with ribosomes. The space between nuclear membranes is called perinuclear. It may contain chloroplasts or leucoplasts, as in heterokonts and cryptophytes. The nuclear matrix contains chromatin, which represents DNA in complex with the main proteins - histones. The exception is the dinophytes, in which the number of histones is small and there is no nucleosomal chromatin organization. The chromatin threads of these algae are arranged in the form of figures of eight. There are from one to several nucleoli in the nucleus, which disappear or persist during mitosis.
Mitosis - indirect division of algae can occur in different ways, but in general the scheme of this process with 4 stages is preserved (Fig. 6).

Rice. 6. Consecutive phases of mitosis: 1 – interphase; 2-4 – prophase; 5 – metaphase; 6– anaphase; 7-9–telophase; 10– cytokinesis
Prophase– the longest phase of mitosis. The most important transformations take place in it: the nucleus increases in volume, instead of a barely noticeable chromatin network, chromosomes appear in it in the form of thin, long, curved and weakly spiraled threads, forming a kind of ball. From the very beginning of prophase, it is clear that chromosomes consist of 2 strands (the result of their replication in interphase). The halves of the chromosomes (chromatids) are located parallel to each other. As prophase develops, the threads become more and more spiralized, and the resulting chromosomes become increasingly shortened and compacted.
At the end of prophase, individual morphological characteristics of chromosomes are revealed. Then the nucleoli disappear, the nuclear membrane fragments into separate short cisterns, indistinguishable from the elements of the EPS, as a result of which the nucleoplasm is mixed with hyaloplasm and myxoplasm is formed; Achromatic filaments—the fission spindle—are formed from the substance of the nucleus and cytoplasm.
The fission spindle is bipolar and consists of bundles of microtubules stretching from one pole to the other. After the destruction of the nuclear membrane, each chromosome is attached to the spindle threads using its centromere. After chromosomes attach to the spindle, they line up in the equatorial plane of the cell so that all centromeres are at the same distance from its poles.
Metaphase. In this phase of mitosis, chromosomes reach maximum compaction and acquire a characteristic shape characteristic of each plant species. Usually they are double-armed, and in these cases, at the point of inflection, called centromere, chromosomes are connected to the achromatin filament of the spindle. In metaphase, it is clearly visible that each chromosome consists of two daughter chromatids. They are located more or less parallel in the equatorial plane of the cell. By the end of the stage, each chromosome is divided into two chromatids, which remain connected only at the centromere. Later, the centromeres also split into two sister ones; while sister centromeres and chromatids face opposite poles.
Anaphase. The shortest phase of mitosis. Daughter chromosomes - chromatids - diverge to opposite poles of the cell. Now the free ends of the chromatids are directed towards the equator, and the kinetochores - towards the poles. It is believed that the chromatids separate due to the contraction of the achromatin spindle filaments, which close to the centromere. Chromosomes become less noticeable due to unwinding and elongation. In the center of the cell (along the equator), sometimes already at this stage fragments of the cell wall - phragmoplast - appear.
Telophase. The process of unwinding continues - despiralization and elongation of chromosomes. Finally, they are lost in the field of view of the optical microscope. The nuclear membrane and nucleolus are restored. The same process occurs as in prophase, only in reverse order. Chromosomes now have one chromatid each. The structure of the interphase nucleus is restored, the spindle changes from a barrel-shaped to a cone-shaped one.
This is how it ends karyotomy– nuclear fission, then comes plasmatomy. Cytoplasmic organelles are distributed between the daughter cells, and some of them (dictyosomes, mitochondria and plastids) undergo significant modifications. Finally it happens cytokinesis– formation of a cell wall between daughter nuclei. From the previous cell two new ones were formed; each of them has a nucleus containing a diploid number of chromosomes.
Depending on the behavior of the nuclear membrane in algae, there are closed, semi-closed And open mitoses. In closed mitosis, chromosome segregation occurs without disruption of the nuclear membrane. In semi-closed mitosis, the nuclear envelope is maintained throughout mitosis, with the exception of the polar zones. In open mitosis, the nuclear membrane disappears in prophase. Depending on the shape of the spindle, divisions are distinguished pleuromitosis And orthomitosis.
In pleuromitosis, a metaphase plate does not form in metaphase and the spindle is represented by two half-spindles located at an angle to each other outside or inside the nucleus. During orthomitosis in metaphase, the chromosomes align with the equator of the bipolar spindle. Depending on the combination of these properties, the following types of mitosis are distinguished in algae (Fig. 7, 8):
Closed extranuclear mitosis

Closed intranuclear mitosis


Semi-closed mitosis


Open mitosis

Rice. 7. Scheme of the main types of mitoses in algae (according to: S.A. Karpov, year). Lines inside or outside the nucleus - spindle microtubules
The centers for organizing microtubules of the mitotic spindle in semi-closed orthomitosis can be kinetosomes and other structures:
– open orthomitosis, found in cryptophytes, goldensea, characeae;
– semi-closed orthomitosis, found in green, red, brown, etc.;
– closed orthomitosis, found in euglenoids;
– closed pleuromitosis, intranuclear or extranuclear, occurs in some dinophytes;
– semi-closed mitosis, during metaphase the centrioles are not at the poles, but in the region of the metaphase plate; can be observed in green trebuxiaceae.



Rice. 8. Diagram comparing (A) closed, (B) metacentric and (C) open mitoses (according to: L.E. Graham, L.W. Wilcox, 2000)
During mitosis, the shape of the spindle and the shape of the spindle poles also vary, as well as the duration of existence of the interzonal spindle. The peak of mitoses occurs during the dark period of the day. In multinucleated cells, nuclear division can occur synchronously. asynchronously, in waves.
Control questions
1. Name the main structural elements of plant cells.
2. The difference in the structure of algae cells from the cells of higher plants.
3. Cell covers of algae.
4. What is theca? In what algae is it found?
5. Main algae pigments. Location of pigments in algae cells.
6. Structure of plastids.
7. Structural features of algae plastids.
8. The structure of mitochondria.
9. Features of the structure of algae mitochondria.
10. Structure of the nucleus and nuclear membranes. Features of nuclear membranes in algae cells.
11. Scheme of mitosis. Characteristics of the phases of mitosis.
12. Types of mitosis in algae cells.
13. What is the difference between pleuromitosis and orthomitosis?
14. Types of algae pseudopodia.
2. GENERAL CHARACTERISTICS OF ALGAE
2.1. Power types
The main type of nutrition in algae is phototrophic type. In all departments of algae there are representatives that are strict (obligate) phototrophs. However, many algae quite easily switch from a phototrophic type of nutrition to the assimilation of organic substances, or heterotrophic food type. However, most often the transition to heterotrophic nutrition in algae does not lead to a complete cessation of photosynthesis, that is, in such cases we can talk about mixotrophic, or mixed type of nutrition.
The ability to grow on organic media in the dark or in light in the absence of carbon dioxide has been shown for many blue-greens, greens, yellow-greens, diatoms, etc. It has been noted that in algae, heterotrophic growth is slower than autotrophic growth in the light.
The diversity and plasticity of algae's feeding methods allows them to have a wide distribution and occupy various ecological niches.
2.2. Types of thalli
The vegetative body of algae is represented thallus, or thallus, not differentiated into organs - root, stem, leaf. Within the thallus structure, algae are distinguished by very large morphological diversity (Fig. 9). They are represented by unicellular, multicellular, and noncellular organisms. Their sizes vary widely: from the smallest single-celled organisms to giant multi-meter organisms. The body shape of algae is also diverse: from the simplest spherical to complexly dissected forms reminiscent of higher plants.
The huge variety of algae can be reduced to several types of morphological structure: monadic, rhizopodial, palmelloid, coccoid, trichal, heterotrichal, parenchymatous, siphonal, siphonocladal.
Monadic (flagellar) type of thallus structure
The most characteristic feature defining this type of structure is the presence of flagella, with the help of which monadic organisms actively move in the aquatic environment (Fig. 9, A). Motile flagellar forms are widespread in algae. Flagellate forms dominate among many groups of algae: euglenophytes, dinophytes, cryptophytes, raphidae, golden algae, and are found in yellow-green and green algae. Brown algae do not have a monadic type of structure in the vegetative state, but monadic stages are formed during reproduction (reproduction). The number of flagella, their length, the nature of placement and movement are varied and have important systematic significance.

Rice. 9. Morphological types of structure of thalli in algae (according to:): A– monadic ( Chlamydomonas); B– amoeboid ( Rhizochrisis); IN– palmelloid ( Hydrurus); G– coccoid ( Pediastrum); D– sarcinoid ( Chlorosarcina); E– filamentous ( Ulotrix); AND– multi-filamentous ( Fritchiela); Z, I– fabric ( Furcellaria, Laminaria); TO– siphonal ( Caulerpa); L– siphonocladal ( Cladophora)
The mobility of monad algae determines the polarity of the structure of their cells and colonies. Flagella are usually attached to the anterior pole of the cell or close to it. The basic shape of the cell is teardrop-shaped with a more or less narrowed anterior flagellar pole. However, monadic organisms often deviate from this basic shape and may be asymmetrical, spiral-shaped, have a tapered posterior end, etc.
The shape of the cell largely depends on the cell covers, which are very diverse (plasmalemma; pellicle; theca; consisting of organic, silica or calcareous scales; house; cell wall). The bizarre outlines of the cells of some golden algae form a kind of intracellular skeleton, consisting of hollow silica tubes. The cell membrane is usually smooth, sometimes bears various projections or is encrusted with iron or calcium salts and then resembles a house. Only small holes are formed in the shell for the exit of flagella.
The polarity of monadic organisms is also manifested in the arrangement of intracellular structures. At the anterior end of the cell there is often a variably arranged pharynx, usually performing an excretory function. Only a few phagotrophic flagellates have a pharynx that functions as a cell mouth - cytostome.
Peculiar organelles characteristic of algae having a monad structure are contractile vacuoles, performing an osmoregulatory function, mucous bodies And stinging structures. Stinging capsules are found in dinophyte, euglenoid, golden, raphidophyte, cryptophyte algae and perform a protective function. The single nucleus occupies a central position in the cells. Chloroplasts, varied in shape and color, can be axial or wall.
The tendency to increase body size is manifested in the formation of various colonies. In the simplest cases, colonies are formed due to nondisjunction of dividing cells. Colonies of ring-shaped, bushy, tree-like, and spherical shapes are observed. Green monadic organisms are mostly characterized by colonies of the type cenobians with a constant number of cells for each type.
Under unfavorable conditions, monadic organisms shed or retract their flagella, losing mobility, and surround themselves with abundant mucus.
The monadic type of structure turned out to be promising. On its basis, other, more complex structures developed.
Rhizopodial (amoeboid) type of structure
The most significant features of the amoeboid type of structure are the absence of strong cell covers and the ability to amoeboid movement, with the help of cytoplasmic outgrowths temporarily formed on the cell surface - pseudopodium. There are several types of pseudopodia, of which algae are most often observed rhizopodia And lodopodia, less often axopodia(Fig. 9, B).
There are no fundamental differences in the structure and mechanism of action of the contractile systems that determine the mobility of monadic and amoeboid organisms at the molecular level. The amoeboid movement probably arose as a result of the adaptation of flagellar cells to simplified living conditions, which led to a simplification of the body structure.
The cells of amoeboid algae contain nuclei, plastids, mitochondria and other organelles characteristic of eukaryotes: contractile vacuoles, stigma and basal bodies capable of forming flagella are often observed.
Many amoeboid organisms lead an attached lifestyle. They can build houses of various shapes and structures: thin, delicate, or rough, thick-walled.
The amoeboid body type is not as widespread as the monadic body type. It is observed only in golden and yellow-green algae.
Palmelloid (hemimonadal) type of structure
Characteristic of this type of structure is the combination of a stationary plant lifestyle with the presence of cellular organelles characteristic of monadic organisms: contractile vacuoles, stigma, flagella. Thus, vegetative cells may have flagella, with the help of which they move within the colonial mucus to a limited extent, or the flagella are preserved in immobile cells in a greatly reduced form.
Cells with the palmelloid (hemimonad) type are characterized by a polar structure. Sometimes the cages are located in houses.
Hemimonad algae often form colonies. In the simplest case, mucus is structureless, and the cells are located inside it in no particular order. Further complexity of such colonies is manifested both in the differentiation of mucus and in a more ordered arrangement of cells within the mucus. Colonies of the dendritic type (genus Hydrurus) (Fig. 9, IN).
The palmelloid (hemimonad) type of structure was an important stage in the morphological evolution of algae in the direction from mobile monadic to typically plant immobile forms.
Coccoid type of structure
This type combines unicellular and colonial algae, immobile in a vegetative state. Cells of the coccoid type are covered with a membrane and have a plant-type protoplast (tonoplast without contractile vacuoles, stigmas, flagella). The loss of signs of a monadic structure in the cell structure in organisms leading a plant-based, sedentary lifestyle, and the acquisition of new structures characteristic of plant cells is the next major step in the evolution of algae according to the plant type.
The huge variety of algae of the coccoid type of structure is associated with the presence of cell covers. The integument determines the presence of various cells: spherical, ovoid, fusiform, ellipsoidal, cylindrical, stellate-lobed, spiral, pear-shaped, etc. The variety of forms is also multiplied thanks to the sculptural decorations of the cell integument - spines, spines, bristles, horny processes.
Coccoid algae form colonies of various shapes, in which the cells are united with or without mucus.
The coccoid type of structure is widespread in almost all divisions of eukaryotic algae (with the exception of Euglenaceae).
In evolutionary terms, the cocoid structure can be considered as the initial one for the emergence of multicellular thalli, as well as siphonal and siphonocladal types of structure (Fig. 9, G, D).
Trichal (filamentous) type of structure
A characteristic feature of the filamentous type of structure is the filamentous arrangement of immobile cells, which are formed vegetatively as a result of cell division occurring predominantly in one plane. Filament cells have a polar structure and can grow only in one direction, coinciding with the axis of the nuclear spindle.
In the simplest cases, thalli of a filamentous structure are composed of cells that are morphologically similar to each other. At the same time, in many algae, in areas of the filaments that become thinner or wider towards the ends, the cells differ in shape from the rest. In this case, often the lower cell, devoid of chloroplasts, turns into a colorless rhizoid or foot. The threads can be simple or branching, single or multi-row, free-living or attached.
The filamentous type of structure is represented among green, red, yellow-green, and golden algae (Fig. 9, E).
Heterotrichal (non-filamentous) type of structure
The heterofilamentous type arose on the basis of the filamentous type. The heterofilamentous thallus consists mostly of horizontal threads creeping along the substrate, performing the function of attachment, and vertical threads rising above the substrate, performing the assimilation function. The latter bear the reproductive organs.
In some algae, vertical filaments are differentiated into internodes And nodes, from which whorls of lateral branches extend, which also have a segmented structure. In addition, additional threads can grow from the nodes, forming a crustal covering of the internodes. The function of attachment to the substrate is performed by colorless rhizoids. This structure can be found in charophytes, green, brown, red, some yellow-green and golden algae (Fig. 9, AND).
Parenchymal (tissue) type of structure
One of the directions in the evolution of the heterofilamentous thallus was associated with the emergence of parenchymatous thalli. The ability for unlimited growth and cell division in different directions led to the formation of voluminous macroscopic thalli with morphofunctional differentiation of cells depending on their position in the thallus (cortex, intermediate layer, pith).
Within this type, there is a gradual complication of thalli from simple plates to complex differentiated thalli with primitive tissues and organs. The parenchymal type of structure is the highest evolutionary stage of morphological differentiation of the algae body. It is widely represented in large algae: brown, red and green - the so-called macrophyte algae (Fig. 10).

Rice. 10. Cross section of the brown algae thallus (by:): 1 – outer bark; 2 – inner cortex; 3 – core
Siphonal type of structure
The siphonal (non-cellular) type of structure is characterized by the absence of cellular partitions inside the thallus, which reaches relatively large, usually macroscopic sizes and a certain degree of differentiation, in the presence of a large number of organelles. Partitions in such a thallus can appear only accidentally, when it is damaged, or during the formation of reproductive organs. In both cases, the process of formation of partitions differs from the formation of a multicellular organism.
The siphonal type of structure is present in some green and yellow-green algae. However, this direction of morphological evolution turned out to be a dead end.
Siphonocladal type of structure
The main feature of the siphonocladal type of structure is the ability to form complexly arranged thalluses, consisting of primarily multinucleated segments, from the primary noncellular thallus. The formation of such a thallus is based on segregation division, in which mitosis does not always end with cytokinesis.
The siphonocladal type of structure is known only in a small group of marine green algae.
2.3. Algae propagation
Reproduction is the main property of living beings. Its essence lies in reproducing its own kind. In algae, reproduction can be carried out asexually, vegetatively and sexually.
Asexual reproduction
Asexual reproduction of algae is carried out using specialized cells - dispute. Sporulation is usually accompanied by division of the protoplast into parts and the release of division products from the shell of the mother cell. Moreover, before the division of the protoplast, processes occur in it leading to its rejuvenation. The release of division products from the shell of the mother cell is the most significant difference between true asexual reproduction and vegetative reproduction. Sometimes only one spore is formed in a cell, but even then it leaves the mother shell.
Spores are usually produced in special cells called sporangia, differing from ordinary vegetative cells in size and shape. They arise as outgrowths of ordinary cells and perform only the function of forming spores. Sometimes spores are formed in cells that do not differ in shape and size from ordinary vegetative cells. Spores also differ from vegetative cells in shape and smaller size. The number of spores in a sporangium ranges from one to several hundred. Spores are a dispersal stage in the algal life cycle.
Depending on the structure there are:
zoospores– motile spores of green and brown algae, may have one, two, four or many flagella, in the latter case the flagella are located in a corolla at the front end of the spore or in pairs over the entire surface;
hemizoospores– zoospores that have lost flagella but retain contractile vacuoles and stigma;
aplanospores– non-motile spores that cover themselves with a membrane inside the mother cell;
motorsports– aplanospores, having the shape of a mother cell;
hypnospores– non-motile spores with thickened shells, designed to survive unfavorable environmental conditions.
In red algae, asexual reproduction occurs using monospore, bispor, tetraspore or polyspore. Monospores do not have a flagellum or membrane. After leaving the mother cell, they are capable of amoeboid movement. Monospores differ from vegetative cells by being ovoid or spherical in shape, rich in nutrients and intensely colored.
The structure of spores and types of sporulation are of great importance for the systematics of algae, since they reflect differences in the organization of the ancestral forms of various groups of algae.
Vegetative propagation
Vegetative propagation in algae can occur in several ways: simple division in two, multiple division, budding, fragmentation of the thallus, stolons, brood buds, paraspores, nodules, akinetes.
Simple division.
This method of reproduction is found only in unicellular forms of algae. Division occurs most simply in cells that have an amoeboid type of body structure.
Division of amoeboid forms. Amoeboid division is possible in any direction. It begins with the elongation of the amoeba's body, and then a partition is outlined at the equator, which divides the body into two more or less equal parts. The division of the cytoplasm is accompanied by division of the nucleus. Sometimes division is preceded by a transition to a stationary state due to the retraction of the legs, and the cell acquires a spherical shape. At the same time, the protoplasm loses its transparency and the contractile vacuole disappears. Towards the end of division, the cell is stretched, laced, and then pseudopods appear.
Division of flagellated forms. In flagellated forms, the most complex types of vegetative propagation occur. Types of reproduction are determined by the level of organization and the degree of cell polarity. In some cryptophyte, golden and green algae, reproduction by simple division in two occurs in a mobile state only along the longitudinal axis and begins from the anterior pole of the cell. In this case, the flagella can go to only one cell or be equally divided between new cells. A cell that does not have a flagellum forms one itself. In most Volvox and Euglena algae, during reproduction, the cell membrane becomes mucus and division occurs in a stationary state. In all flagellated forms that have a shell, the cells are divided into two equal or unequal parts. After separation, the old shell is shed and a new one is formed.
Division of coccoid forms. In algae with a coccoid type of cell structure, vegetative reproduction acquires the typical features of division of a stationary plant cell with a well-defined cell wall. In its simplicity, it approaches the amoeboid type of vegetative reproduction and is carried out by simple division of the cell in two.
Budding.
Cells of filamentous branched algae are characterized by two ways of vegetative reproduction: simple division in two and budding. The combination of these methods of reproduction causes lateral branching of filamentous algae.
Fragmentation.
Fragmentation is inherent in all groups of multicellular algae and manifests itself in different forms: the formation of hormogonium, regeneration of detached parts of the thallus, spontaneous loss of branches, regrowth of rhizoids. The cause of fragmentation may be mechanical factors (waves, currents, animal gnawing), or the death of some cells. An example of the latter method of fragmentation is the formation of hormogonia in blue-green algae. Each hormogonium can give rise to a new individual. Reproduction by parts of thalli, characteristic of red and brown algae, does not always lead to the resumption of normal plants. Seaweed growing on rocks and rocks is often partially or completely destroyed by wave action. Their detached fragments or entire thalli are not able to re-attach themselves to solid soils due to the constant movement of water. In addition, the attachment organs are not formed again. If such thalli find themselves in quiet places with a muddy or sandy bottom, they continue to grow while lying on the ground. Over time, the older parts die off and the branches extending from them turn into independent thalli; in such cases they speak of unattached, or free-living, forms of the corresponding species. The algae change greatly: their branches become thinner, narrower and branch weaker. Unattached forms of algae do not form organs of sexual and asexual reproduction and can only reproduce vegetatively.
Reproduction by shoots, stolons, brood buds, nodules, akinetes.
In the tissue forms of green, brown and red algae, vegetative reproduction takes on its complete form, which differs little from the vegetative reproduction of higher plants. While retaining the ability to regenerate parts of the thallus, tissue forms acquire specialized formations that perform the function of vegetative propagation. Many species of brown, red, green and chara algae have shoots on which new thalli grow. On the thalli of some brown and red algae, brood buds (propagules) develop, which fall off and grow into new thalli.
With the help of unicellular or multicellular overwintering nodules, seasonal renewal of charophytic algae occurs. Some filamentous algae (for example, green ulothrix algae) reproduce by akinetes - specialized cells with a thickened shell and a large amount of reserve nutrients. They are able to survive adverse conditions.
Sexual reproduction
Sexual reproduction in algae is associated with the sexual process, which consists of the fusion of two cells, resulting in the formation of a zygote that grows into a new individual or produces zoospores.
There are several types of sexual reproduction in algae:
hologamy(conjugation) – without the formation of specialized cells;
gametogamy– with the help of specialized cells – gametes.
Hologamy. In the simplest case, the process occurs by the fusion of two immobile vegetative cells lacking cell membranes. In unicellular flagellated forms of algae, the sexual process is carried out by the fusion of two individuals.
When the contents of two flagellated vegetative cells merge, the sexual process is called conjugation. During conjugation, the fusion of two cells occurs, which perform the function of germ cells - gametes. The fusion of cell contents occurs through a specially formed conjugation channel, resulting in a zygote, which is subsequently covered with a thick membrane and turns into a zygospore. If the rate of flow of cell contents is the same, a zygote is formed in the conjugation channel. In this case, the division of cells into male and female is conditional.
Gametogamy. Sexual reproduction in algae, including unicellular algae, most often occurs by dividing the contents of the cells and the formation of specialized germ cells in them - gametes. In all green and brown algae, male gametes have flagella, but female gametes do not always have them. In primitive algae, gametes are formed in vegetative cells. In more highly organized forms, gametes are located in special cells called gametangia. A vegetative cell or gametangium can contain from one to several hundred gametes. Depending on the size of the merging gametes, several types of gametogamy are distinguished: isogamy, heterogamy, oogamy.
If the merging gametes have the same shape and size, this sexual process is called isogamy.
If the merging gametes have the same shape, but different sizes (the female gamete is larger than the male one), then they speak of heterogamy.
A sexual process in which an immobile large cell merges - egg and a mobile small male cell - sperm, called oogamy. Gametangia with eggs are called archegonia or oogonia, and with spermatozoa – antheridia. Male and female gametes can develop on the same individual (monoecious) or on different individuals (dioecious). The zygote formed as a result of the fusion of gametes, after some changes, turns into a zygospore. The latter is usually covered with a dense shell. Zygospore can remain dormant for a long time (up to several months) or germinate without a dormant period.
Autogamy. A special type of sexual process. It consists in the fact that the cell nucleus divides meiotically, of the four nuclei formed, two are destroyed, and the remaining two nuclei merge, forming a zygote, which, without a rest period, increases in size and turns into an auxospore. This is how individuals rejuvenate.
2.4. Life cycles of algae
Life cycle, or development cycle, is a set of all stages of development of organisms, as a result of which, from certain individuals or their rudiments, new individuals and rudiments similar to them are formed. The stage of aging, leading to the death of the individual, and periods of rest extend beyond the life cycle. The development cycle can be simple or complex, which is associated with the ratio of diploid and haploid nuclear phases, or forms of development(Fig. 11).

Rice. 11. Life cycles of algae (according to:): I – haplobiont with zygotic reduction; II – haplodiplobiont with sporic reduction; III – diplobiont with gametic reduction; IV – haplodiplobiont with somatic reduction. The dominant phase in cases I and III is multicellular; if it is unicellular, then it is long-lasting and capable of mitotic reproduction; 1 – haploid phase; 2 – diploid phase
The concept of life cycle is associated with the alternation of generations. Under generation understand the totality of individuals considered in relation to ancestors and descendants living at a close time, and genetically related to it.
A simple life cycle is characteristic of cyanobacteria, in which sexual reproduction is not found. Their life cycles are complete ( big) And small. The small life cycle corresponds to certain branches of the large cycle and leads to the repeated formation of intermediate age states of cyanobacteria individuals . The development cycle of cyanobacteria, thus, includes certain segments of the development of one or a number of successive generations of a specific systematic form: from the primordium of an individual to the emergence of new primordia of the same type.
In most algae with a sexual process, depending on the time of year and external conditions, different forms of reproduction are observed (sexual and asexual), with a change in haploid and diploid nuclear phases. The changes undergone by an individual between the same phases of development constitute his life cycle.
Organs of sexual and asexual reproduction can develop on the same individual or on different individuals. Plants that produce spores are called sporophytes, and the forming gametes are gametophytes. Plants that can produce both spores and gametes are called gametosporophytes. Gametosporophytes are characteristic of many algae: green (Ulvacaceae), brown (Ectocarpaceae) and red (Bangieaceae). The development of reproductive organs of one type or another is determined by the temperature of the environment. For example, on the lamellar thalli of red algae Porphyra Tenera at temperatures below 15–17 °C, organs of sexual reproduction are formed, and at higher temperatures, organs of asexual reproduction are formed. In general, in many algae, gametes develop at a lower temperature than spores. The development of certain reproductive organs is also influenced by other factors: light intensity, day length, chemical composition of water, including its salinity.
Gametophytes, gametosporophytes and sporophytes of algae may not differ in appearance or have well-defined morphological differences. Distinguish isomorphic(similar) and heteromorphic(different) changes in forms of development, which are identified with the alternation of generations. In most gametosporophytes, alternation of generations does not occur. Sometimes gametophytes and sporophytes, without differing morphologically, exist in different environmental conditions; in some cases they differ morphologically. For example, in red algae Porphyra Tenera sporophytes have the form of branching single-row filaments that are embedded in the calcareous substrate (mollusk shells, rocks). They grow preferentially in low light and penetrate the substrate to great depths. The gametophytes of this algae have the form of plates and grow in good light near the water's edge and in the tidal zone.
With heteromorphic alternation of generations, the structure of sporophytes and gametophytes differs in some cases quite significantly. Thus, in green algae from the genera Acrosiphony And Spongomorpha the gametophyte is multicellular, several centimeters high, and the sporophyte is unicellular, microscopic. Other ratios of gametophyte and sporophyte sizes are also possible. In brown algae Sugars the gametophyte is microscopic, and the sporophyte is up to 12 m long. In most algae, gametophytes and sporophytes are independent plants. In a number of species of red algae, sporophytes grow on gametophytes, and in some brown algae, gametophytes develop inside the sporophyte thallus.
A heteromorphic change in developmental forms, when a clearly defined separation of the sporophyte from the gametophyte is observed, is characteristic of more highly organized groups of algae. In this case, one of the forms, most often the gametophyte, is microscopic. It is believed that the heteromorphic development cycle of algae arose from an isomorphic one. The methods of development of the gametophyte and sporophyte are of great importance in the taxonomy of algae. The most complex and diverse development cycles, not found in other algae, are characteristic of red algae.
Change of nuclear phases.
During the sexual process, as a result of the fusion of gametes and their nuclei, the number of chromosomes in the nucleus doubles. At a certain stage of the development cycle, during meiosis, the number of chromosomes is reduced, as a result of which the resulting nuclei receive a single set of chromosomes. The sporophytes of many algae are diploid, and meiosis in their development cycle coincides with the moment of formation of spores, from which haploid gametosporophytes or gametophytes develop. This meiosis is called sporic reduction. Sporophytes of more primitive red algae (genera Cladophora, Ectocarpus and many others) along with haploid spores form diploid spores, which again develop into sporophytes. Spores appearing on gametosporophytes serve for self-renewal of mother plants. Sporophytes and gametophytes of algae at the highest stages of evolution strictly alternate without self-renewal.
In a number of algae, meiosis occurs in the zygote. This meiosis is called zygotic reduction and is found in a number of species of green and charophyte algae. In freshwater volvox and ulothrix algae, the sporophyte is represented by a unicellular zygote, which produces up to 32 zoospores, the mass of which is many times greater than the parent gametes, i.e. essentially a sporic reduction is observed.
Some groups of algae have gametic reduction, which is characteristic of animals, and not of plant organisms. In these algae, meiosis occurs during the formation of gametes, while the remaining cells of the thallus remain diploid. Such a change in nuclear phases is inherent in diatoms and brown fucus algae, which are widespread throughout the globe (which include the most widespread species of marine algae), and among green algae, in the large genus Cladophora. Development with gametic reduction of the nucleus is believed to give these algae certain advantages over others.
If reduction division occurs in sporangia before the formation of spores of asexual reproduction (sporic reduction), then there is an alternation of generations - a diploid sporophyte and a haploid gametophyte. This type of life cycle is called haplobiont with sporic reduction. It is characteristic of some green algae, many brown and red algae.
Finally, in a few algae, meiosis occurs in the vegetative cells of the diploid thallus (somatic reduction), from which haploid thalli then develop. Such life cycle with somatic reduction known from red and green algae.
Control questions
Types of algae nutrition.
Types of algae thallus.
Characteristics of the monadic morphological structure.
Characteristics of rhizopodial morphological structure. Types of cytoplasmic processes.
Characteristics of palmelloid morphological structure.
Characteristics of coccoid morphological structure.
Characteristics of the trichal morphological structure.
Characteristics of heterotrichal morphological structure.
Characteristics of parenchymal morphological structure.
Characteristics of the siphonal morphological structure.
Characteristics of siphonocladal morphological structure.
12. Asexual reproduction. Types of disputes.
13. Types of vegetative propagation of algae.
14. Types of sexual reproduction of algae.
15. How do sporophytes and gametophytes differ?
16. What is heteromorphic and isomorphic change of generations?
17. Change of nuclear phases in the life cycle of algae. Sporical, zygotic and gametic reduction.
3. ECOLOGICAL GROUPS OF ALGAE
Algae are distributed throughout the globe and are found in various aquatic, terrestrial and soil biotopes. Various ecological groups are known: algae of aquatic habitats, terrestrial algae, soil algae, algae of hot springs, algae of snow and ice, algae of hypersaline springs.
3.1. Algae of aquatic habitats
3.1.1. Phytoplankton
The term “phytoplankton” means a collection of plant organisms floating in the water column. Planktonic algae are the main, and in some cases the only, producer of primary organic matter, on the basis of which all life in a body of water exists. The productivity of phytoplankton depends on a complex of various factors.
Planktonic algae live in a variety of bodies of water - from the ocean to a puddle. Moreover, the greater diversity of environmental conditions in inland water bodies compared to the seas also determines a significantly greater diversity of species composition and ecological complexes of freshwater plankton.
Phytoplankton of freshwater ecosystems characterized by a clearly defined seasonality. In each season, one or several groups of algae predominate in a reservoir, and during periods of intensive development, often only one species dominates. So in winter, under the ice (especially when the ice is covered with snow), phytoplankton is very poor or almost absent due to lack of light. The vegetative development of plankton algae as a community begins in March - April, when the level of sunlight becomes sufficient for photosynthesis of algae even under ice. At this time, quite numerous small flagellates appear - euglenophytes, dinophytes, golden ones, as well as cold-loving diatoms. During the period of ice breaking up before temperature stratification is established, which usually happens when the upper layer of water is heated to 10–12C°, the rapid development of the cold-loving complex of diatoms begins. In summer, when the water temperature is above 15C°, the maximum productivity of blue-green, euglenoid and green algae is observed. Depending on the trophic and limnological type of the reservoir, water “blooming” may occur at this time, caused by the development of blue-green and green algae.
One of the significant features of freshwater phytoplankton is the abundance of temporary planktonic algae in it. A number of species, which are generally considered to be typically planktonic, in ponds and lakes have a bottom or periphyton (attached to some object) phase in their development.
Marine phytoplankton consists mainly of diatoms and dinophytes. Of the diatoms, representatives of the genera are especially numerous Chaetoceros, Rhizosolenia, Thalassiosira and some others absent from freshwater plankton. The composition of flagellar forms of dinophyte algae in marine phytoplankton is very diverse. Representatives of primnesiophytes are very numerous in marine phytoplankton; they are represented in fresh waters by only a few species. Although the marine environment is relatively homogeneous over large areas, similar homogeneity is not observed in the distribution of marine phytoplankton. Differences in species composition and abundance are often pronounced even in relatively small areas of sea water, but they are especially clearly reflected in the large-scale geographic zonality of distribution. Here the ecological effect of the main environmental factors is manifested: water salinity, temperature, light and nutrient content.
Planktonic algae usually have special adaptations for living in suspension. Some have various kinds of outgrowths and body appendages - spines, bristles, horny outgrowths, membranes. Others form colonies that secrete mucus abundantly. Still others accumulate substances in their bodies that increase their buoyancy (droplets of fat in diatoms, gas vacuoles in blue-greens). These formations are much more developed in marine phytoplankters than in freshwater ones. One of the adaptations for existing suspended in the water column is the small body size of planktonic algae.
3.1.2. Phytobenthos
Phytobenthos refers to a set of plant organisms that are adapted to exist in an attached or unattached state at the bottom of reservoirs and on various objects, living and dead organisms in the water.
The possibility of benthic algae growing in specific habitats is determined by both abiotic and biotic factors. Among biotic factors, competition with other algae and the presence of consumers play a significant role. This leads to the fact that certain types of benthic algae do not grow at all depths and not in all water bodies with suitable light and hydrochemical conditions. Light is especially important for the growth of benthic algae as photosynthetic organisms. But the degree of its use is influenced by other environmental factors: temperature, content of biogenic and biologically active substances, oxygen and inorganic carbon sources, and most importantly, the rate of entry of these substances into the thallus, which depends on the concentration of substances and the speed of water movement. As a rule, places with intense water movement are characterized by lush development of benthic algae.
Benthic algae growing in active conditions water movement, gain advantages over algae growing in sedentary waters. The same level of photosynthesis can be achieved by phytobenthos organisms under flow conditions with less light, which promotes the growth of larger thalli. The movement of water, moreover, prevents the settling of silt particles on rocks and stones, which interfere with the fixation of algae buds, favor the growth of bottom algae, washing away algae-eating animals from the soil surface. Finally, although during strong currents or strong surf the algae thalli are damaged or torn from the ground, the movement of water still does not prevent the settlement of microscopic species of algae or microscopic stages of macrophyte algae.
The influence of water movement on the development of benthic algae is especially noticeable in rivers, streams and mountain streams. In these reservoirs there is a group of benthic organisms that prefer places with strong currents. In lakes where there are no strong currents, wave motion becomes of primary importance. In the seas, waves also have a significant impact on the life of benthic algae, in particular on their vertical distribution.
In the northern seas, the distribution and abundance of benthic algae is influenced by ice. Depending on its thickness, movement and hummocking, algae thickets can be destroyed (erased) to a depth of several meters. Therefore, for example, in the Arctic, perennial brown algae ( Fucus, Laminaria) is easiest to find near the shore among boulders and rock ledges that impede the movement of ice.
The life of benthic algae is influenced in many ways by temperature. Along with other factors, it determines their growth rate, the pace and direction of development, the moment of formation of their reproductive organs, and the geographic zonality of distribution.
Intensive development of algae is also facilitated by moderate content in water. nutrients. In fresh waters, such conditions are created in shallow ponds, in the coastal zone of lakes, in river backwaters, in the seas - in small bays.
If in such places there is sufficient lighting, hard soils and weak water movement, then optimal conditions for the life of phytobenthos are created. In the absence of water movement and its insufficient enrichment with nutrients, benthic algae grow poorly. Such conditions exist in rocky bays with a large bottom slope and significant depths in the center, since nutrients from bottom sediments are not carried into the upper horizons. In addition, macroscopic seaweeds, which serve as substrates for many small forms of benthic algae, may be absent in such habitats.
Sources of nutrients in water are coastal runoff and bottom sediments. The role of the latter is especially great as accumulators of organic residues. In bottom sediments, as a result of the vital activity of bacteria and fungi, mineralization of organic residues occurs; complex organic substances are converted into simple inorganic compounds available for use by photosynthetic plants.
In addition to light, water movement, temperature and nutrient content, the growth of benthic algae depends on presence of herbivorous aquatic animals– sea urchins, gastropods, crustaceans, fish. This is especially noticeable in the thickets of kelp algae, which are large in size. In tropical seas, in some places, fish completely eat up green, brown and red algae with a soft thallus. Gastropods, crawling along the bottom, eat microscopic algae and small seedlings of macroscopic species.
The predominant bottom algae of continental water bodies are diatoms, green, blue-green and yellow-green filamentous algae, attached or not attached to the substrate.
The main benthic algae of the seas and oceans are brown and red, sometimes green, macroscopic attached thallous forms. All of them can be overgrown with small diatoms, blue-green and other algae.
Depending on the place of growth, the following ecological groups are distinguished among benthic algae:
epiliths– grow on the surface of hard soil (rocks, stones);
epipelites– inhabit the surface of loose soils (sand, silt);
epiphytes– epizoites– live on the surface of plants/animals;
endophytes– endozoites or endosymbionts– live inside the body of plants/animals, but feed independently (have chloroplasts and photosynthesize);
endoliths– live in calcareous substrate (rocks, mollusk shells, crustacean shells).
Sometimes a group of organisms is isolated fouling, or periphyton. Organisms included in this group live on objects that are mostly moving or flown around by water. In addition, they are removed from the bottom and are exposed to different light, food and temperature conditions than truly bottom-dwelling organisms.
The fouling composition includes microalgae and macrophyte algae. Microscopic algae (blue-green and diatoms) form a mucous bacterial-algal-detrital film on a substrate introduced into the aquatic environment. Then macroalgae (red, brown and green) settle on the primary microfilm along with the animals. This creates serious interference in human economic activity. Due to fouling, the speed of ships and the efficiency of hydroacoustic devices decreases, fuel consumption increases, and underwater structures become heavier and corrode. In addition, the slimy film formed by fouling can disrupt the operation of water pipes, clog the openings of water intakes and pipelines, and disrupt heat exchange processes in refrigeration units.
Attached fouling organisms that live on underwater structures in the intertidal zone and at depths of up to 1 m are usually eliminated in winter by prolonged drying and abrasion by ice. Therefore, every year in the spring-summer period, fouling communities are formed here, characteristic of the pioneer stage of biological succession. The dominant species of such communities, along with barnacles and mollusks, are often macrophyte algae. In the sublittoral zone of underwater structures - from a depth of 0.7-0.9 m to their base (6-12 m) - perennial fouling develops. Its composition is dominated by brown algae from the genera Saccharina And Costaria. The biomass of these large algae in temperate latitudes can be very significant, amounting to tens of kilograms per square meter.
Fouling algae can also exist in the air ( aerophyton). Of these, green and blue-green algae predominate. Under certain conditions, aerophyton algae can damage industrial and building materials, architectural monuments, paintings, etc., if they are not protected by toxic coatings. The cause of damage is the metabolic products of fouling agents, mainly organic acids. Aerophyton algae are especially common in the humid tropics, where there is enough heat, moisture and dust of organic origin, which is a breeding ground for their development. Biodamage from them can be significant.
Epilites. This group includes attached algae. They populate the surface of stones, forming crust-like coverings or flat pads, or have special attachment organs - rhizoids. Intensive development of epiliths is observed in reservoirs with a hard bottom and fast flowing water. Typical epiliths are representatives of golden algae from the genus Hydrurus, brown algae from the genera Saccharina, Kelp, Costaria and etc.
Epipelites. Loose algae spreading along the bottom, binding and strengthening the substrate. They are often represented by diatoms, aureus, euglenoids, cryptophytes, and dinophytes freely crawling on the substrate. The attachment organs of epipelites are sometimes short rhizoids that cannot take deep root. Only charophyte algae with their long rhizoids develop well on the muddy bottom.
Usually, the organs of attachment of epiliths and epipeliths are special formations - sole, leg, foot, mucous cord or mucous pad, cushion, etc.
Epiphytes/epizoites. Algae use living organisms as a substrate. Epizoites are algae that settle on animals. On the surface of mollusk shells there are small green ( Edogonium, Cladophora, Ulva) and red ( Gelidium, Palmaria,) seaweed; on sponges - green, blue-green and diatoms. Epizoites live on crustaceans, rotifers, less commonly on aquatic non-insects or larvae, worms and even larger animals. Epizoites include species of green and charophyte algae from the genera Chlorangiella, Charatiochloris, Korzhikoviella, Chlorangiopsis etc. Most epizoites cannot exist in isolation from the substrate. Algae usually die on dead animals or on their shells shed during molting.
Epiphytes are algae that live on plants. Short-term connections arise between the substrate plant (basiphyte) and the epiphyte plant. The complex and interesting phenomenon of epiphytism is still poorly understood. There are frequent cases of double or even triple epiphytism, when some algae settling on other, larger forms themselves are a substrate for other, smaller or microscopic species. Sometimes the physiological state of the substrate plant is important for the development of epiphytes. The number of epiphytes, as a rule, increases as the basifite algae ages. For example, the greatest species richness of epiphytic aedogonia algae is observed on dead aquatic plants ( Manna, Reed, Sedge).
Endophytes/endozoites, or endosymbionts
Endosymbionts, or intracellular symbionts - algae that live in the tissues or cells of other organisms (invertebrate animals or algae). They form a kind of ecological group. Intracellular symbionts do not lose the ability to photosynthesize and reproduce inside host cells. A variety of algae can be endosymbionts, but the most numerous are endosymbioses of unicellular green and yellow-green algae with unicellular animals. The algae participating in such symbioses are respectively called zoochlorella And zooxanthellae. Green and yellow-green algae form endosymbioses with multicellular organisms: sponges, hydras, etc. Endosymbioses of blue-green algae with protozoa are called syncyanoses. Often, other types of cyanobacteria can settle in the mucus of some blue-green species. They usually use ready-made organic compounds, which are formed in abundance during the breakdown of the mucus of the colony of the host plant, and multiply intensively.
The most common endophytes are representatives of goldens (species of the genera Chromulina, Myxochloris) and green (genus Chlorochitrium, Chlamydomyx) algae that settle in the body of duckweed and sphagnum mosses. Green algae genus Carteria settles in the epidermal cells of the ciliated worm Convolute, one species of the genus Chlorella– in the vacuoles of protozoa, and species of the genus Chlorococcum– in the cells of cryptophyte algae Cyanophora.
3.1.3. Algae of extreme aquatic ecosystems
Hot spring algae. Algae that grow at temperatures of 35–85 °C are called thermophilic. Often, high environmental temperatures are combined with a high content of mineral salts or organic substances (heavily contaminated hot wastewater from factories, factories, power plants or nuclear plants). Typical inhabitants of hot waters are blue-green algae and, to a lesser extent, diatoms and green algae.
Algae of snow and ice. Algae that grow on the surface of ice and snow are called cryophilic. When developing in large numbers, they can cause green, yellow, blue, red, brown or black “blooming” of snow or ice. Among cryophilic algae, green, blue-green and diatom algae predominate. Only a few of these algae have dormant stages; most lack any special morphological adaptations to tolerate low temperatures.
Algae from salt water bodies got the name halophilic or halobionts. Such algae grow at high salt concentrations in water, reaching 285 g/l in lakes with a predominance of table salt and 347 g/l in Glauberian lakes. As salinity increases, the number of algae species decreases; Only a few of them can tolerate very high salinity. In oversaline (hyperhaline) water bodies, unicellular mobile green algae predominate - hyperhalobes, whose cells lack a membrane and are surrounded by plasmalemma ( Asteromonas, Pedinomonas). They are distinguished by an increased content of sodium chloride in the protoplasm, high intracellular osmotic pressure, and the accumulation of carotenoids and glycerol in the cells. In some settled reservoirs, such algae can cause red or green “blooming” of the water. The bottom of hyperhaline reservoirs is sometimes completely covered with blue-green algae; Among them, species from the genera predominate Oscillatorium, Spirulina etc. With a decrease in salinity, an increase in the species diversity of algae is observed: in addition to blue-green algae, diatoms appear (species of the genera Navicula, Nietzsche).
3.2. Algae of non-aquatic habitats
Although the main living environment for most algae is water, due to the eurytopic nature of this group of organisms, they successfully colonize a variety of out-of-water habitats. In the presence of at least periodic moisture, many of them develop on various ground objects - rocks, tree bark, fences, etc. A favorable habitat for algae is soil. In addition, communities of endolith algae are also known, the main living environment of which is the surrounding calcareous substrate.
Communities formed by algae in extra-aquatic habitats are divided into aerophilic, edaphilic and lithophilic.
3.2.1. Aerophilic algae
The main living environment of aerophilic algae is the air surrounding them. Typical habitats are the surface of various extra-soil hard substrates (rocks, stones, tree bark, house walls, etc.). Depending on the degree of moisture, they are divided into two groups: air and water-air. Air algae They live in conditions of only atmospheric moisture and experience a constant change in humidity and drying. Water-air algae are exposed to constant irrigation with water (under the spray of waterfalls, in the surf zone, etc.).
The living conditions of these algae are very peculiar and are characterized, first of all, by the frequent change of two factors - humidity and temperature. Algae that live in conditions of exclusively atmospheric moisture are forced to frequently transition from a state of excessive moisture (for example, after a rainstorm) to a state of minimal moisture during dry periods, when they dry out so much that they can be ground into powder. Aquatic algae live in conditions of relatively constant moisture, however, they also experience significant fluctuations in this factor. For example, algae that live on rocks irrigated by the spray of waterfalls experience a moisture deficit in the summer, when the flow decreases significantly. Aerophilic communities are also susceptible to constant temperature fluctuations. They get very hot during the day, cool down at night, and freeze in winter. True, some aerophilic algae live in fairly constant conditions (on the walls of greenhouses). But in general, relatively few algae, represented by microscopic unicellular, colonial and filamentous forms of blue-green and green algae and, to a much lesser extent, diatoms, have adapted to the unfavorable conditions of existence of this group. Aerophilic forms are also known among red algae of the genus Porphyridium and etc.; they are found on stones and old walls of greenhouses. The number of species found in aerophilic groups approaches 300. When aerophilic algae develop in mass quantities, they usually take the form of powdery or slimy deposits, felt-like masses, soft or hard films and crusts.
On the bark of trees, the usual settlers are the ubiquitous green algae from the genera Pleurococcus, Chlorella, Chlorococcus. Blue-green algae and diatoms are found much less frequently on trees. There is evidence that predominantly green algae grow on gymnosperms.
The systematic composition of algae groups living on the surface of exposed rocks is different. Diatoms and some, mostly unicellular, green algae develop here, but representatives of blue-green algae are most common in these habitats. Algae and accompanying bacteria form “mountain tan” (rock films and crusts) on crystalline rocks of various mountain ranges. The debris that accumulates in rock recesses is usually inhabited by single-celled green algae and blue-green algae. Algae growths are especially abundant on the surface of wet rocks. They form films and growths of various colors. As a rule, species equipped with thick mucous membranes live here. Depending on the light intensity, the mucus can be colored more or less intensely, which determines the color of the growths. They can be bright green, golden, brown, purple, almost black, depending on the species that form them. Particularly characteristic of irrigated rocks are representatives of blue-green algae, such as species of the genera Gleocapsa, Tolipothrix, Spirogyra etc. In the growths on wet rocks you can also find diatoms from the genera Frustulia, Akhnantes and etc.
Thus, aerophilic algae communities are very diverse and arise both under completely favorable and extreme conditions. External and internal adaptations to this lifestyle are varied and similar to the adaptations of soil algae, especially those developing on the soil surface.
3.2.2. Edaphilic algae
The main living environment of edaphophilic algae is soil. Typical habitats are the surface and thickness of the soil layer, which has a physical and chemical effect on the bionts. Depending on the location of algae and their lifestyle, three groups are distinguished within this type: terrestrial algae, massively developing on the soil surface under atmospheric moisture conditions; water-terrestrial seaweed, developing en masse on the surface of the soil, constantly saturated with water; soil algae, inhabiting the thickness of the soil layer.
Soil as a biotope is similar to both aquatic and aerial habitats: it contains air, but it is saturated with water vapor, which ensures breathing with atmospheric air without the threat of drying out. Property. which fundamentally distinguishes the soil from the above-mentioned biotopes is its opacity. This factor has a decisive influence on the development of algae. However, in the soil thickness, where light does not penetrate, viable algae are found at a depth of up to 2 m in virgin lands and up to 2.7 m in arable lands. This is explained by the ability of some algae to switch to heterotrophic nutrition in the dark.
A small number of algae are found in the deep layers of the soil. To maintain their viability, soil algae must have the ability to tolerate unstable humidity, sudden temperature fluctuations and strong insolation. These properties are ensured by a number of morphological and physiological features. For example, it has been noted that soil algae are relatively small in size compared to the corresponding aquatic forms of the same species. As cell size decreases, their water-holding capacity and resistance to drought increase. An important role in the drought resistance of soil algae is played by the ability to produce abundant mucus - slimy colonies, covers and wrappers consisting of hydrophilic polysaccharides. Due to the presence of mucus, algae quickly absorb water when moistened and store it, slowing down drying. Soil algae stored in an air-dry state in soil samples demonstrate amazing viability. If such soil is placed on a nutrient medium after decades, it will be possible to observe the development of algae.
A characteristic feature of soil algae is the “ephemerality” of the growing season - the ability to quickly move from a state of dormancy to active life and vice versa. They are also able to withstand temperature fluctuations over a very wide range: from -200 to +84°C. Soil algae (mostly blue-green) are resistant to ultraviolet and radioactive radiation.
The vast majority of soil algae are microscopic forms, but they can often be seen on the soil surface with the naked eye. The massive development of such algae can cause greening of the slopes of ravines and the sides of forest roads.
In terms of systematic composition, soil algae are quite diverse. Among them, blue-green and green algae are represented in approximately equal proportions. Yellow-green algae and diatoms are less diverse in the soils.
3.2.3. Lithophilic algae
The main living environment of lithophilic algae is the opaque dense calcareous substrate surrounding them. Typical habitats are deep within hard rocks of a certain chemical composition, surrounded by air or submerged in water. There are two groups of lithophilic algae: drilling algae, which actively penetrate into the limestone substrate; tuff-forming algae, depositing lime around their body and living in the peripheral layers of the environment they deposit, within the limits accessible to water and light. As sediment builds up, it dies off.
Control questions
1. Describe the main ecological groups of algae in aquatic habitats: phytoplankton and phytobenthos.
2. Differences between freshwater and marine phytoplakton. Representatives of marine and freshwater phytoplankton.
3. Morphological adaptations of algae to the planktonic lifestyle.
4. Seasonal changes in qualitative and quantitative indicators of freshwater phytoplankton.
5. Differences between freshwater and marine phytobenthos. Systematic composition of marine and freshwater phytobenthos.
6. Ecological groups of phytobenthos in relation to the substrate (epilites, epipelites, epiphytes, endophytes).
7. What is fouling? What algae can make up this ecological group?
8. Aerophilic algae. Adaptations to extreme environmental conditions. Systematic composition of aerial algae.
9. Edaphilic algae. Adaptations to environmental conditions. Systematic composition of soil algae.
10. Lithophilic algae.
4. ROLE OF ALGAE IN NATURE AND PRACTICAL IMPORTANCE
The role of algae in natural ecosystems. In aquatic biocenoses, algae play the role of producers. Using light energy, they are able to synthesize organic substances from inorganic ones. According to radiocarbon dating, the average primary production of the oceans due to the vital activity of algae is 550 kg of carbon per 1 hectare per year. The total value of its primary production is 550.2 billion tons (in raw biomass) per year and, according to scientists, the contribution of algae to the total production of organic carbon on our planet ranges from 26 to 90%. Algae play an important role in the nitrogen cycle. They are able to use both organic (urea, amino acids, amides) and inorganic (ammonium and nitrate ions) sources of nitrogen. A unique group are blue-green algae, which are capable of fixing nitrogen gas, converting it into compounds available to other plants.
Algae - oxygen producers. Algae, in the process of their life activity, release oxygen necessary for the respiration of aquatic organisms. In the aquatic environment (especially in the seas and oceans), algae are practically the only producers of free oxygen. In addition, they play a large role in the overall oxygen balance on Earth, since the oceans serve as the main regulator of the oxygen balance in the Earth's atmosphere.
Algae are a medium for other aquatic organisms. By forming underwater forests, macrophyte algae create highly productive ecosystems that provide food, shelter and protection to many other living organisms. It was established that a column of water with a volume of 5 liters containing one specimen of brown algae Cystoseira contains up to 60 thousand individuals of various invertebrate animals, including mollusks, mites and crustaceans.
Algae - pioneers of vegetation. Terrestrial algae can settle on bare rocks, sand and other barren places. After they die, the first layer of future soil is formed. Soil algae participate in the processes of formation of soil structure and fertility.
Algae as a geological factor. The development of algae in past geological epochs has led to the formation of a number of rocks. Together with animals, algae took part in the formation of reefs in the oceans. Settled closer to the surface of the water, they formed the ridges of these reefs. Reef structures of red algae are known in the Crimea as the peaks of Yayla and others. Blue-green algae participated in the formation of stromatolite limestones, chara algae - in the formation of charocyte limestones (similar deposits were found in Tuva). Cocolithophores take part in the formation of Cretaceous rocks (Cretaceous rocks are 95% composed of the remains of the shells of these algae). The massive accumulation of diatom shells led to the formation of diatomite (mountain flour), large deposits of which were discovered in the Primorsky Territory, the Urals and Sakhalin. Algae were the starting material for liquid and solid petroleum-like compounds - sapropels, hot shale, coal.
The active activity of algae in the formation of rocks has been noted in some regions at the present time. They absorb calcium carbonate and form mineralized products. These processes are especially active in tropical waters with high temperatures and low partial pressure.
Boring algae are of greatest importance in the destruction of rocks. They slowly and persistently loosen calcareous substrates, making them available for weathering, crumbling and erosion.
Symbiotic relationships with other organisms. Algae form several important symbioses. Firstly, they form lichens with fungi, and secondly, as zooxants they live together with some invertebrate animals, such as sponges, ascidians, and reef corals. A number of cyanophytes form associations with higher plants.
Algae are of great practical importance in everyday life and human economic activity, bringing both benefit and harm. Large, mainly seaweeds have been known since ancient times and have long been used in human agriculture.
Algae as a food product. Humans eat mainly seaweed; they are especially widely used by residents of Southeast Asia and the Pacific Islands. In China, the use of algae in the diet has been known since the 9th century BC. e. Among macrophyte algae (multicellular green, brown and red) there are no poisonous species, because they do not contain alkaloids - substances with a narcotic and poisonous effect. About 160 species of different algae are used for food. In terms of nutritional qualities, algae are not inferior to many agricultural crops. They contain a large percentage of protein, carbohydrates and fats. Algae are an excellent source of vitamins C, A, D, group B, riboflavin, pantothenic and folic acids, and microelements.
Of microscopic algae, blue-green terrestrial species of the genus are used as food. Nostok, which serve as food in China and South America. In Japan, they eat barley bread "tengu" - these are thick layers of dense gelatinous mass on the slopes of some volcanoes, consisting of blue-green algae from the genera Gleocapsa, Geoteke, Microcystis with an admixture of bacteria. Spirulina used by the Aztecs back in the 16th century, preparing cakes from dried seaweed, and the population of the Lake Chad region in North America still prepares a product called dihe from this seaweed. Spirulina contains high amounts of protein and is widely cultivated in a number of countries.
Algae as fertilizer. Algae contain a sufficient amount of organic and mineral substances, so they have long been used as fertilizers. The advantages of such fertilizers are that they do not contain weed seeds and spores of phytopathogenic fungi, and their potassium content is superior to almost all types of fertilizers used. Nitrogen-fixing blue-green algae are widely used in rice fields instead of nitrogen fertilizers. It has been shown that algae fertilizers can increase seed germination, yield, and disease resistance.
Medicinal properties of algae. Algae are widely used in folk medicine as an anthelmintic and for the treatment of a number of diseases, such as goiter, nervous disorders, sclerosis, rheumatism, rickets, etc. It has been shown that extracts of many types of algae contain antibiotic substances and can reduce blood pressure. Extracts from Sargassum, Kelp and Saharina in experiments on mice, they suppressed the growth of sarcoma and leukemic cells. In the USA and Japan, drugs have been obtained from them that help remove radionuclides from the body. The efficiency of such sorbents reaches 90–95%.
Algae as a source of industrial raw materials. Since the last century, algae have been used to produce soda and iodine. Currently, alginic acid and its salts - alginates, as well as carrageenans and agar are obtained from algae.
Mannitol alcohol is obtained from brown algae - a necessary raw material for the pharmaceutical and food industries in the manufacture of medicines and food products for diabetics.
Negative role of algae. A number of algae (blue-green, dinophyte, golden, green) produce toxins that can cause various diseases in animals, plants and humans, some of which can be fatal. Among the dinophyte algae that cause “red tides” in vast marine areas, the species of the genera are toxic Gymnodinium, Noctiluca, Amphidinium etc. The largest number of toxic species was identified among blue-green algae. The action of blue-green algae toxins is several times greater than that of poisons such as curare and botulin. The toxicity of algae manifests itself in the mass death of aquatic organisms, waterfowl, poisoning and other diseases of people that occur through inhalation, use of water, consumption of shellfish, fish, etc.
With strong development - “blooming of water bodies”, some algae (golden, yellow-green, blue-green) can give the water an unpleasant odor and taste, making the water unsuitable for drinking.
Excessive algae growth can prevent water from passing through the filters of water intake structures. It is known that algae fouling of ships significantly increases operating costs. Macrophytes can contribute to the corrosion of materials on oil platforms and other underwater marine structures.
The problem of fouling is perhaps the oldest problem in ocean exploration. Any object that comes into contact with the marine environment is soon covered with a mass of organisms attached to it: animals and algae. The total area of submerged substrates is about 20% of the surface area of the upper shelf. The total biomass of fouling amounts to millions of tons, the damage from it amounts to billions of dollars (Zvyagintsev, 2005). In the biological aspect, this is a natural process that forms an integral part of the life of the hydrosphere. At the same time, the phenomenon of fouling suggested to man the idea of growing a number of valuable species of mollusks in marine fisheries on an industrial scale ( Oysters, Mussels, Scallops, Pearl mussels) and algae ( Saccharines, Porphyry, Gracilaria, Euchema and etc.). Algae are pioneer fouling organisms. Microalgae, together with bacteria, form a primary microfilm on the surface of artificial substrates added to water, which serves as a substrate for the sedimentation of other hydrobionts. Macroalgae, together with crustaceans, mollusks, hydroids and other animals, often form the initial stages of perennial fouling communities.
Control questions
1. The role of algae in increasing land fertility.
2. The role of algae in aquatic ecosystems.
3. The role of algae in terrestrial ecosystems.
4. The importance of algae in geological processes.
5. Nutritional and biological value of algae. What seaweed can be eaten?
6. Medicinal properties of algae.
7. Why is the growth of golden and yellow-green algae in reservoirs undesirable? What is “blooming” of water bodies?
8. Algae that cause poisoning of animals and humans.
9. Fouling phenomenon. The role of algae in fouling communities.
5. MODERN SYSTEMATICS OF ALGAE
The classification of living organisms has occupied the minds of people since the time of Aristotle. The Swedish botanist Carl Linnaeus was the first to apply the name Algae to a group of plants in the 18th century and began phycology(from Greek phycos – algae and logos – teaching) as a science. Among algae, Linnaeus distinguished only four genera: Chara, Fucus, Ulva and Conferva. In the 19th century, the majority (several thousand) of modern algae genera were described. The large number of new genera necessitated grouping them into higher-ranking taxa. Initial attempts at classification were based solely on external features of the thallus. The first to propose the color of algae thallus as a fundamental character for the establishment of large taxonomic groups, or megataxa, was the English scientist W. Harvey (Harvey, 1836). He identified large series: Chlorospermeae - green algae, Melanospermeae - brown algae and Rhodospermeae - red algae. They were later renamed Chlorophyceae, Phaeophyceae, and Rhodophyceae, respectively.
The foundations of modern algae taxonomy were laid in the first half of the 20th century by the Czech scientist A. Pascher. He established 10 classes of algae: Blue-green, Red, Green, Golden, Yellow-green, Diatom, Brown, Dinophyte, Cryptophyte and Euglenaceae. Each class is characterized by a specific set of pigments, reserve products and the structure of flagella. These constant differences between large taxa prompted us to consider them as independent phylogenetic groups, unrelated, and to abandon the concept of algae - Algae as a specific taxonomic unit.
Thus, the word “algae” is actually not a systematic, but an ecological concept and literally means “what grows in water.” Algae are lower plants that contain the vast majority of chlorophyll, are capable of phototrophic nutrition and live primarily in water. All algae, except charophytes, unlike higher plants, do not have multicellular reproductive organs with covers of sterile cells.
Modern systems differ mainly in the number and volume of megataxa - divisions and kingdoms. The number of departments varies from 4 to 10-12. In Russian phycological literature, almost each of the above classes corresponds to a department. In foreign literature, there is a tendency towards consolidation of departments and, accordingly, a decrease in their number.
The most common in classification schemes is Parker's scheme (Parker, 1982). It recognizes the division between prokaryotic and eukaryotic forms. Prokaryotic forms do not have membrane-surrounded organelles in their cells. Prokaryotes include Bacteria and Cyanophyta (Cyanobacteria). Eukaryotic forms include all other algae and plants. The algae division has long been a subject of debate. Harvey (1836) divided algae primarily by color. Although many more divisions are now recognized, the composition of pigments, biochemical and structural features of the cell structure are given great importance. P. Silva (1982) distinguishes 16 main classes. The classes differ in pigmentation, storage products, cell wall features and ultrastructure of flagella, nucleus, chloroplasts, pyrenoids and ocelli.
New information on the ultrastructure of algae, obtained in recent decades using the methods of electron microscopy, genetics and molecular biology, makes it possible to study the smallest details of the cell structure. “Explosions” of information periodically prompt scientists to reconsider established traditional ideas about the taxonomy of algae. The constant flow of new information stimulates new approaches to classification, and each proposed scheme inevitably remains approximate. According to modern data, organisms traditionally considered among lower plants go beyond the scope of the Plant Kingdom. They are included in a large number of independently evolving groups. The table shows megataxa, which include algae, in different interpretations. As can be seen, different algal taxa can be found in different phyla; the same phyla can unite different ecological and trophic groups of organisms (table).
More than 100 years ago K.A. Timiryazev perspicaciously noted that “there is neither plant nor animal, but there is one inseparable organic world. Plants and animals are only average values, only typical ideas that we form, abstracting from the known characteristics of organisms, attaching exceptional importance to some, neglecting others.” Now we can't help but admire his amazing biological intuition.
The modern algae system outlined in this textbook includes 9 divisions: Blue-green, Red, Diatoms, Heterokonts, Haptophytes, Cryptophytes, Dinophytes, Green, Charophytes and Euglenophytes. The similarity in the composition of pigments, the structure of the photosynthetic apparatus and flagella served as the basis for uniting the classes of algae with a golden-brown color into one large group - Heterokontae, or heteroflagellate algae (Ochrophyta).
Megasystem of organisms classified as lower plants
|
Empire |
Kingdom |
Department (Type) |
Trophogroup |
|
Eubacteria/Prokaryota |
Cyanobacteria/Bacteria |
Cyanophyta/ Cyanobacteria |
Seaweed |
|
Excavates/Eukaryota |
Euglenobiontes/ Protozoa |
Euglenophyta/ Euglenozoa Acrasiomycota |
Seaweed Myxomycetes |
|
Rhizaria/Eukaryota |
Cercozoa/ Plantae |
Chlorarachniophyta Plasmodiophoromy-cota |
Algae Myxomycetes |
|
Rhizaria/Eukaryota |
Myxogasteromycota Dictyosteliomycota |
Myxomycetes Myxomycetes |
|
|
Choromalveola-tes/ Eukaryota |
Straminopilae/ Chromista/ Heterokontobiontes |
Labyrinthulomycota -Oomycota Heterokontophyta |
Myxomycetes Fungi Algae |
|
Choromalveola-tes/ Eukaryota |
Haptophytes/ Chromista |
Prymnesiophyta/Haptophyta |
Seaweed |
|
Choromalveola-tes/ Eukaryota |
Cryptophytes/ Chromista |
Seaweed |
|
|
Choromalveola-tes/ Eukaryota |
Alveolates/Protozoa |
Dinophyta/Myzozoa |
Seaweed |
|
Plantae/ Eukaryota |
Glaucophytes/ Plantae |
Glaucocystophyta/ Glaucophyta |
Seaweed |
|
Plantae/ Eukaryota |
Rhodobiontes/ Plantae |
Cyanidiophyta Rhodophyta |
Algae Algae |
|
Plantae/ Eukaryota |
Chlorobiontes/ Plantae |
Chlorophyta Charophyta |
Algae Algae |
Algae, unicellular and multicellular forms of benthic algae. All morphological types of thallus are found here, except for rhizopodial unicellular and large multicellular forms with a complex structure. Many filamentous green algae attach to the substrate only in the early stages of development, then they become free-living, forming mats or balls.
| Green algae | ||||||||||||
| Diversity of siphon algae. Illustration from Ernst Haeckel's book Kunstformen der Natur, 1904 |
||||||||||||
| Scientific classification | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| International scientific name | ||||||||||||
|
Chlorophyta Pascher, 1914 |
||||||||||||
|
||||||||||||
Green algae are the most extensive department of algae at this time: according to rough estimates, this includes from 13,000 to 20,000 species. All of them are distinguished primarily by the pure green color of their thalli, similar to the color of higher plants and caused by the predominance of chlorophyll over other pigments.
Structure
The flagellar cells of green algae are isokont - the flagella have a similar structure, although they can vary in length. There are usually two flagella, but there can also be four or many. Flagella of green algae do not have mastigonemes (unlike heterokonts), but may have graceful hairs or scales.
Life cycles
The life cycles of green algae are very diverse. There are all kinds of types here.
Haplobiont with zygotic reduction ( Hydrodictyon reticulatum, Eudorina). Biflagellate gametes are released from the parent cell through a pore in its membrane; the fusion of gametes is carried out using a tube. Next, the zygote turns into a resting zygospore, and after a period of physiological dormancy it germinates to form 4 zoospores (as a result of meiotic division). Each zoospore forms a polyhedron and germinates, forming small spherical networks of adherent zoospores.
Haplo-diplobiont with sporic reduction ( Ulva, Ulothrix, some types Cladophora). Biflagellate isogametes emerge from the mother cell, after which the gametes formed by different filaments merge in water. A four-flagellate zygote is formed, which actively floats in the water. After this, it descends onto some substrate and becomes covered with a dense shell, thus turning into a club-shaped cell (codiolum), followed by a stage of physiological rest. When favorable conditions occur, it germinates into 4-16 zoospores or aplanospores, which, after a short period of swimming, attach to the substrate and grow into new filaments. Various factors activate the exit from the dormant state: an increase in temperature, a change in the environment, etc.
Diplobiont with gametic reduction ( Bryopsis). The planozygote settles and grows into a filamentous thallus with a large nucleus; the nucleus divides, thus forming stephanokont zoospores that grow into the vegetative thallus.
Especially a lot of green algae develops in the spring, when all the stones in the littoral zone are covered with a continuous emerald coating of green algae, sharply contrasting with the white snow lying on the coastal stones. A fleecy green carpet on the stones is formed by developing filaments - ulotrix ( Ulothrix) and urospora ( Urospora). In summer, a lot of Egagropyla often develops ( Aegagropila linnaei) (syn. Cladophora aegagropila), which often looks like a green mucous mass. On the open rocky coast, bright green branched bushes form acrosiphony ( Acrosiphonia).
Role in nature and use
Some green algae (such as ulva) are widely eaten. Chlorella is used as an indicator of the level of water pollution and is contained on spaceships and submarines to purify the air from carbon dioxide.
Today, green algae are considered the most extensive group, which has about 20 thousand species. This includes both unicellular organisms and colonial forms, as well as plants with large multicellular thallus. There are representatives that live in water (sea and fresh), as well as organisms adapted to survive on land in conditions of high humidity.
Department Green algae: brief description
The main distinguishing feature of representatives of this group is their coloring - all species are characterized by green or green-yellow coloring. This is due to the main pigment of cells - chlorophyll.
As already mentioned, the department brings together completely different representatives. There are unicellular and colonial forms, as well as multicellular organisms with a large, differentiated thallus. Some unicellular representatives move with the help of flagella; multicellular ones, as a rule, are attached to the bottom or live in the water column.
Although there are organisms with naked cells, most representatives possess a cell wall. The main structural component of the cell membrane is cellulose, which, by the way, is considered an important systematic characteristic.
The number, size and shape of chloroplasts in a cell may vary depending on the type of plant. The main pigment is chlorophyll, in particular the a and b forms. As for carotenoids, plastids contain mainly beta-carotene and lutein, as well as small amounts of neosanthin, zeaxanthin and violaxanthin. Interestingly, the cells of some organisms have an intense yellow or even orange color - this is due to the accumulation of carotenes outside the chloroplast.
Some unicellular green algae have a specific structure - an eye, which reacts to light in the blue and green spectrum.
The main storage product is starch, the granules of which are contained mainly in plastids. Only some representatives of the order have reserve substances deposited in the cytoplasm.
Department Green algae: methods of reproduction
In fact, representatives of this order are characterized by almost all possible methods of reproduction. can occur through (unicellular representatives without a cell membrane), fragmentation of the thallus (this method is typical for multicellular and colonial forms). In some species, specific nodules are formed.
Asexual reproduction is represented by the following forms:
- zoospores - cells with flagella, capable of active movement;
- aplanospores - such spores do not have a flagellar apparatus, but well-developed cells are not capable of active movement;
- autospores - this type of spores is primarily associated with adaptation to the external environment. In this form, the body can wait out dry conditions and other unfavorable conditions.
Sexual reproduction can also be diverse - this includes oogamy, heterogamy, hologamy, as well as isogamy and conjugation.
Order Green algae: characteristics of some representatives
This group includes many famous representatives of the plant world. For example, spirogyra and chlorella are also included in the order.
Chlamydomonas is a fairly well-known genus of green algae, which is of great practical importance. This group includes single-celled organisms with a red eye and a large chromatophore that contains pigments. It is Chlamydomonas that causes the “blooming” of ponds, puddles and aquariums. In the presence of sunlight, organic matter is produced through photosynthesis. But this organism can absorb substances from the external environment. Therefore, chlamydomonas is often used to purify water.
According to their characteristics (color, structure, methods of reproduction, etc.), algae are divided into several types (divisions): green, blue-green, golden, diatoms, yellow-green, pyrrophytic, euglenic, red and brown algae.
Let's give a brief description of the types.
Green algae - Chlorophyta
Unicellular, colonial, multicellular and noncellular forms. Multicellular forms are represented mainly by filamentous algae. Some are distinguished by a complex internal structure, reminiscent of higher plants in appearance.
Algae are purely green in color, however, in addition to green chlorophyll, the chromatophores contain yellow pigments - carotene and xanthophyll. The cell membrane consists of fiber. Chromatophores with pyrenoids.
Reproduction is carried out by vegetative, asexual and sexual means. Vegetative propagation occurs by dividing the organism into parts. Asexual reproduction is carried out by motile zoospores with flagella of equal size (usually 2-4 of them) or aplaiospores - immobile spores.
With the help of zoospores, green algae not only reproduce, but also disperse. The sexual process of reproduction is varied. Representatives of green algae are Chlamydomonas, Spirogyra, Chlorella, Ulothrpx, Cladophora, Closterium, etc.
Blue-green algae - Cyanophyta
Unicellular, colonial and filamentous forms. Algae have blue-green, yellow-green, olive green and other types of colors. The color is explained by the presence of four pigments in blue-green algae: green chlorophyll, blue phycocyan, red phycoerythryp-pa and yellow carotene. These algae do not have a formed chromatophore and nucleus, flagellar stages and the sexual process are absent. Blue-green algae include: oscillatoria, nostoc, gleotrichia, anabena, etc.
Golden algae - Chrysophyta
Single-celled and colonial forms. They contain chlorophyll and phycochrysipus, which is why the color of representatives of this group of algae is golden or brownish-yellow. The cells are in some cases naked or covered with a poorly differentiated protoplasmic membrane; golden algae are found, the body of which is dressed in a shell or enclosed in a house.
Some forms are mobile and move with the help of flagella, while other forms are immobile in a vegetative state. They reproduce by division or zoospores.
Capable of forming cysts to withstand unfavorable conditions. The sexual process is very rare. Representatives of this type of algae are mallomonas, dinobrion, chrysameba, etc.
Diatoms - Bacillariophyta
Unicellular and colonial organisms with a silicified shell consisting of two halves called valves. Chromatophore I<ел-того или светло-бурого цвета от наличия в нем, кроме хлорофилла, бурого пигмента диатомина. Размножение осуществляется путем деления клеток на две, у некоторых диатомовых наблюдается образование двужгутиковых зооспор. Известен половой процесс. К диатомовым водорослям относятся пинну л я р ия, навикула, плевросигма, гомфонема, сиредра, мелозира и др.
Yellow-green, or heteroflagellate, algae - Xanthophyta, or Heterocontae
These include unicellular, colonial, filamentous and noncellular forms. These algae contain, in addition to chlorophyll, yellow pigments - xanthophyll and carotene; their color varies from light to dark yellow-green. Reproduction occurs by longitudinal cell division, zoospores (they are characterized by the presence of two flagella of unequal size and unequal structure), and autospores. The sexual process is known. Representative: botridium.
Pyrrophyte algae - Pyrrophyta
Unicellular and colonial forms. In addition to chlorophyll, algae contain the pigment pyrrophyll, which gives the algae a brown and brownish-yellow color. The cells are naked or covered with armored membranes. They reproduce by division, zoosiors, autosiors. They form cysts. Sexual reproduction is rare. Pyrrophyte algae include: peridinium, ceracium, etc.
Euglenophyta - Euglenophyta
Unicellular motile forms with one or two flagella, sometimes without them; the cells are bare, the role of the shell is played by the outer layer of protoplasm, sometimes the cell is located in the house. Most algae are green in color, sometimes light green due to the presence of xaptophylla. Reproduction occurs by longitudinal division, the sexual process is unknown. Representatives of euglena algae are euglena and facus.
Red algae, or purple algae, Rhodophyta
They live mainly in the seas, only a few live in fresh waters. These are multicellular algae, colored red.
(with different shades). The color of algae is associated with the presence in them, in addition to chlorophyll, of other pigments - phycoerythrin and phycocyan.
Asexual reproduction is carried out by aplano-spores. The sexual process is very complex and is characterized by the presence of male organs - antheridia and female organs - oogonia, or carpogones.
The representative is batrachospermum.
Brown algae, Phaeophyta
The name was given due to the yellow-brown color of the thallus, caused by the presence, in addition to green chlorophylls, of a large number of brown carotenoid pigments. Multicellular, predominantly macroscopic algae (the largest organism living in water is the brown algae macrocystis, which reaches a length of 60 m, growing by 45 cm per day).
Reproduction is vegetative, asexual and sexual. Gametes and zoospores bear two flagella on the side, different in length and morphology. Brown algae are widespread in all seas of the planet; they often form underwater forests, reaching their greatest development in the seas of temperate and subpolar latitudes, where they are the main source of organic matter in the coastal zone. In tropical latitudes, the largest accumulation of brown algae is in the Sargasso Sea. Representatives of only a few genera live in desalinated and fresh waters, for example, Pleurocladia, Streblonema, Lithoderma.